

Time-resolved neutron scattering provides new insight into protein substrate processing by a AAA+ unfoldase
- PMID: 28102317
- PMCID: PMC5244417
- DOI: 10.1038/srep40948
Time-resolved neutron scattering provides new insight into protein substrate processing by a AAA+ unfoldase
Abstract
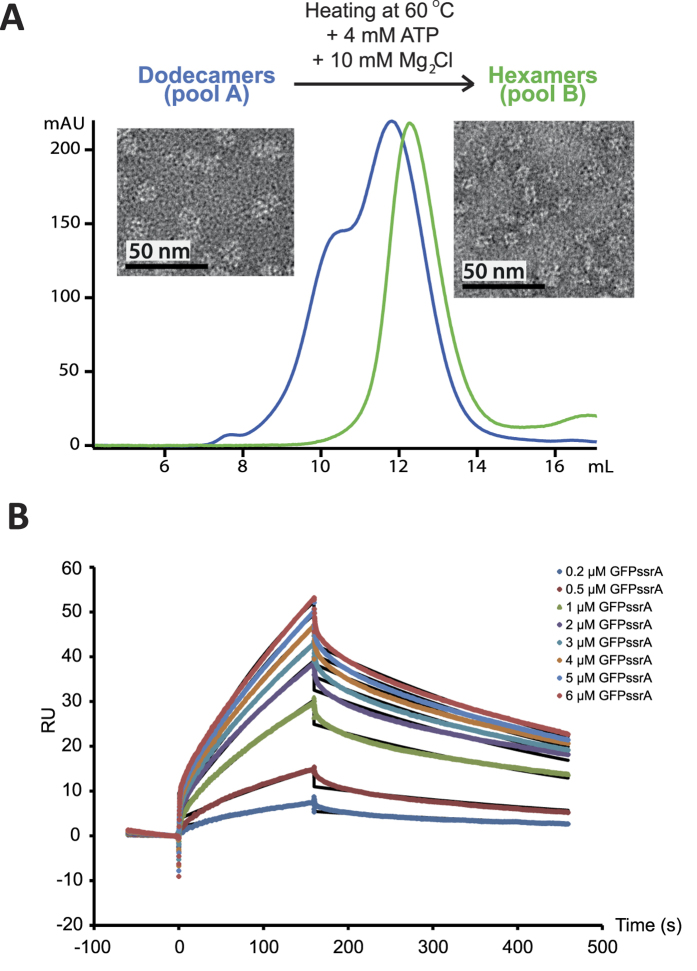
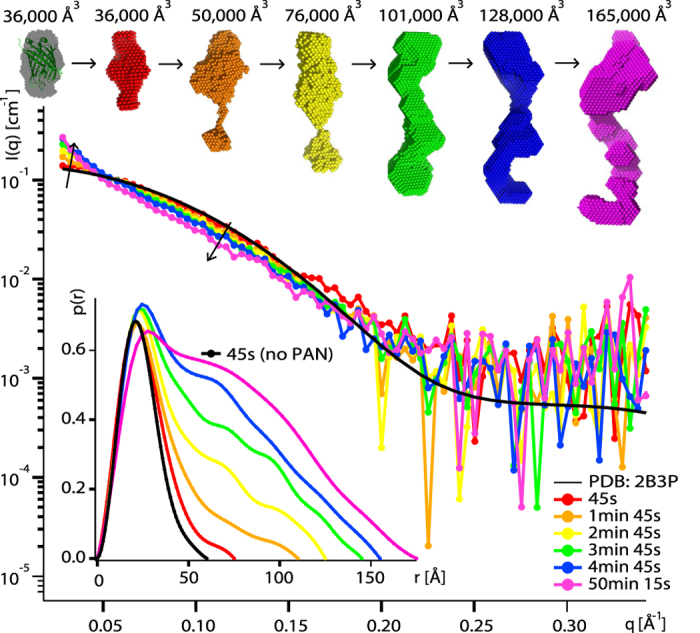
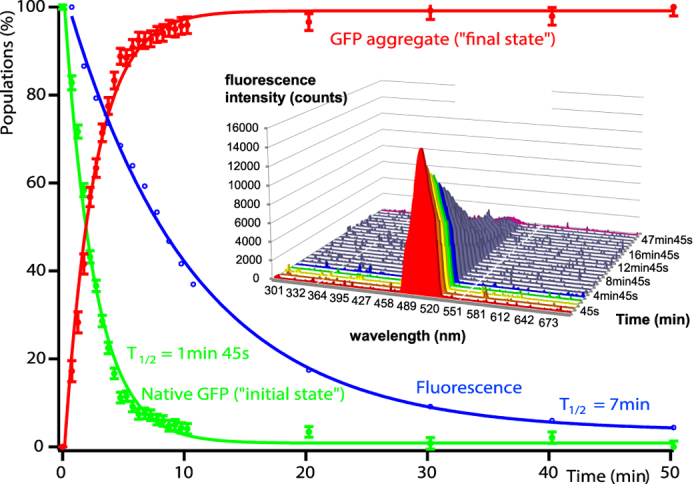
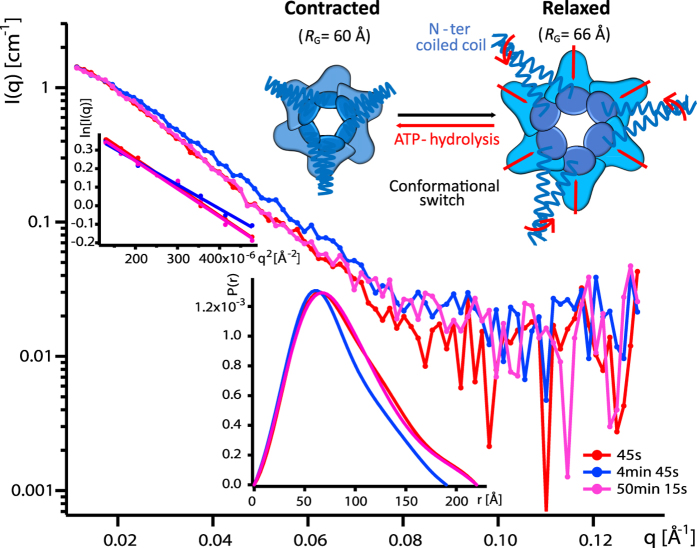
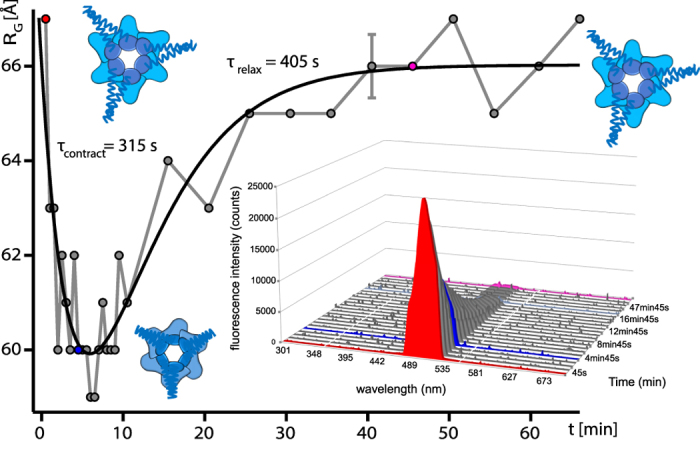
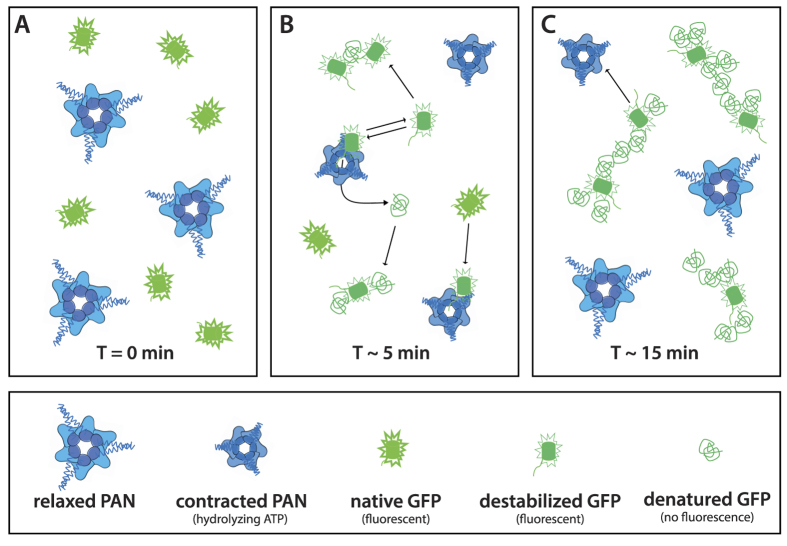
We present a combination of small-angle neutron scattering, deuterium labelling and contrast variation, temperature activation and fluorescence spectroscopy as a novel approach to obtain time-resolved, structural data individually from macromolecular complexes and their substrates during active biochemical reactions. The approach allowed us to monitor the mechanical unfolding of a green fluorescent protein model substrate by the archaeal AAA+ PAN unfoldase on the sub-minute time scale. Concomitant with the unfolding of its substrate, the PAN complex underwent an energy-dependent transition from a relaxed to a contracted conformation, followed by a slower expansion to its initial state at the end of the reaction. The results support a model in which AAA ATPases unfold their substrates in a reversible power stroke mechanism involving several subunits and demonstrate the general utility of this time-resolved approach for studying the structural molecular kinetics of multiple protein remodelling complexes and their substrates on the sub-minute time scale.
Figures
Similar articles
-
Proteasomes: unfoldase-assisted protein degradation machines.Biol Chem. 2019 Dec 18;401(1):183-199. doi: 10.1515/hsz-2019-0344. Biol Chem. 2019. PMID: 31665105 Review.
-
Structures and mechanisms of AAA+ protein complexes in DNA processing.Curr Opin Struct Biol. 2025 Jun;92:103056. doi: 10.1016/j.sbi.2025.103056. Epub 2025 May 6. Curr Opin Struct Biol. 2025. PMID: 40334521 Review.
-
Overcoming fluorescence loss in mEOS-based AAA+ unfoldase reporters through covalent linkage.Protein Expr Purif. 2025 Aug;232:106724. doi: 10.1016/j.pep.2025.106724. Epub 2025 Apr 28. Protein Expr Purif. 2025. PMID: 40306474
-
A real-time analysis of GFP unfolding by the AAA+ unfoldase PAN.J Magn Reson. 2023 May;350:107431. doi: 10.1016/j.jmr.2023.107431. Epub 2023 Apr 5. J Magn Reson. 2023. PMID: 37058954
-
Bromodomain AAA+ ATPases get into shape.Nucleus. 2020 Dec;11(1):32-34. doi: 10.1080/19491034.2020.1741304. Nucleus. 2020. PMID: 32191554 Free PMC article. Review.
Cited by
-
Hiding the Elephant in the Room with Experimental Neutrons.Biophys J. 2020 Jul 21;119(2):234-235. doi: 10.1016/j.bpj.2020.05.038. Epub 2020 Jun 24. Biophys J. 2020. PMID: 32640187 Free PMC article. No abstract available.
-
Deuteration Aiming for Neutron Scattering.Biophys Physicobiol. 2021 Feb 6;18:16-27. doi: 10.2142/biophysico.bppb-v18.003. eCollection 2021. Biophys Physicobiol. 2021. PMID: 33954079 Free PMC article.
-
Recent Progress in Solution Structure Studies of Photosynthetic Proteins Using Small-Angle Scattering Methods.Molecules. 2023 Nov 3;28(21):7414. doi: 10.3390/molecules28217414. Molecules. 2023. PMID: 37959833 Free PMC article. Review.
-
Examining Membrane Proteins by Neutron Scattering.Methods Mol Biol. 2020;2168:147-175. doi: 10.1007/978-1-0716-0724-4_7. Methods Mol Biol. 2020. PMID: 33582991
-
Observing Protein Degradation by the PAN-20S Proteasome by Time-Resolved Neutron Scattering.Biophys J. 2020 Jul 21;119(2):375-388. doi: 10.1016/j.bpj.2020.06.015. Epub 2020 Jun 24. Biophys J. 2020. PMID: 32640186 Free PMC article.
References
-
- Sauer R. T. & Baker T. A. AAA+ Proteases: ATP-Fueled Machines of Protein Destruction. Annu Rev Biochem 80, 587–612 (2011). - PubMed
-
- Prakash S. & Matouschek A. Protein unfolding in the cell. TRENDS Biochem Sci 29, 593–600 (2004). - PubMed
-
- Baumeister W., Walz J., Zuhl F. & Seemuller E. The Proteasome: Paradigm of a Self-Compartmentalizing Protease. Cell 92, 367–380 (1998). - PubMed
-
- Ciechanover A. Proteolysis: from the lysosome to ubiquitin and the proteasome. Nat Rev Mol Cell Biol 6, 79–86 (2005). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
