

Rfx2 Stabilizes Foxj1 Binding at Chromatin Loops to Enable Multiciliated Cell Gene Expression
- PMID: 28103240
- PMCID: PMC5245798
- DOI: 10.1371/journal.pgen.1006538
Rfx2 Stabilizes Foxj1 Binding at Chromatin Loops to Enable Multiciliated Cell Gene Expression
Abstract
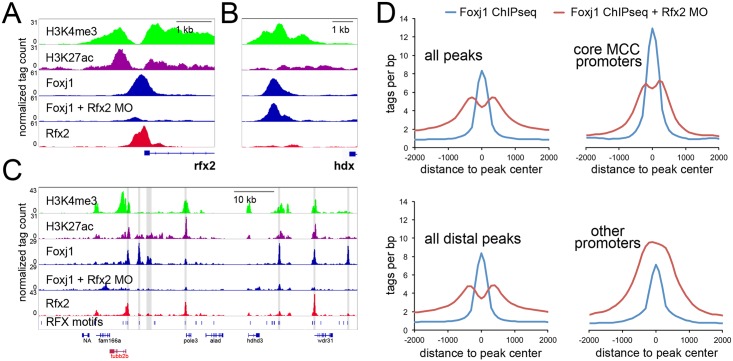
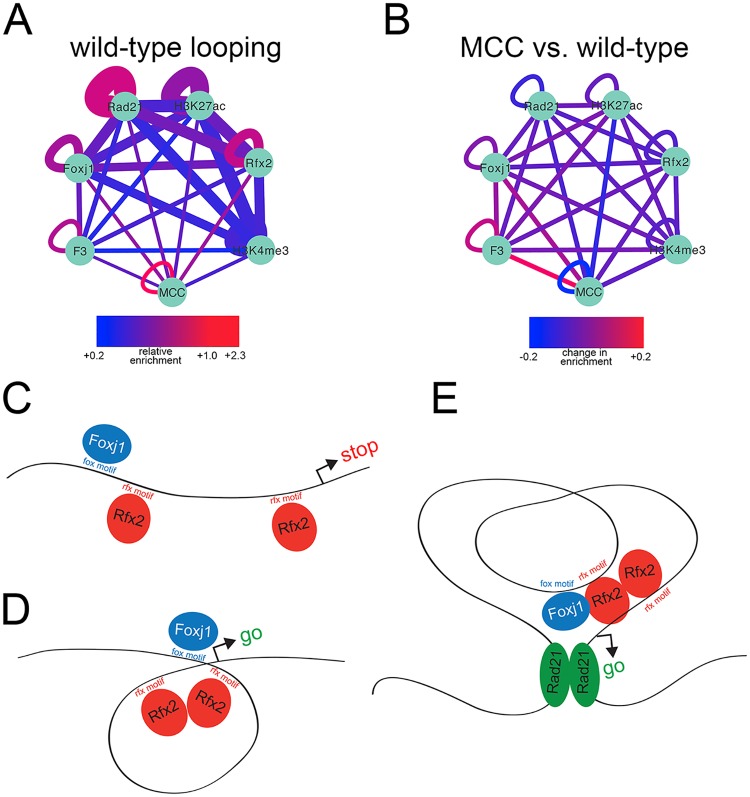
Cooperative transcription factor binding at cis-regulatory sites in the genome drives robust eukaryotic gene expression, and many such sites must be coordinated to produce coherent transcriptional programs. The transcriptional program leading to motile cilia formation requires members of the DNA-binding forkhead (Fox) and Rfx transcription factor families and these factors co-localize to cilia gene promoters, but it is not clear how many cilia genes are regulated by these two factors, whether these factors act directly or indirectly, or how these factors act with specificity in the context of a 3-dimensional genome. Here, we use genome-wide approaches to show that cilia genes reside at the boundaries of topological domains and that these areas have low enhancer density. We show that the transcription factors Foxj1 and Rfx2 binding occurs in the promoters of more cilia genes than other known cilia transcription factors and that while Rfx2 binds directly to promoters and enhancers equally, Foxj1 prefers direct binding to enhancers and is stabilized at promoters by Rfx2. Finally, we show that Rfx2 and Foxj1 lie at the anchor endpoints of chromatin loops, suggesting that target genes are activated when Foxj1 bound at distal sites is recruited via a loop created by Rfx2 binding at both sites. We speculate that the primary function of Rfx2 is to stabilize distal enhancers with proximal promoters by operating as a scaffolding factor, bringing key regulatory domains bound by Foxj1 into close physical proximity and enabling coordinated cilia gene expression.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Gualdi R, Bossard P, Zheng M, Hamada Y, Coleman JR, Zaret KS. Hepatic specification of the gut endoderm in vitro: cell signaling and transcriptional control. Genes Dev. 1996;10: 1670–82. Available: http://www.ncbi.nlm.nih.gov/pubmed/8682297 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
