

Aryl hydrocarbon receptor/cytochrome P450 1A1 pathway mediates breast cancer stem cells expansion through PTEN inhibition and β-Catenin and Akt activation
- PMID: 28103884
- PMCID: PMC5244521
- DOI: 10.1186/s12943-016-0570-y
Aryl hydrocarbon receptor/cytochrome P450 1A1 pathway mediates breast cancer stem cells expansion through PTEN inhibition and β-Catenin and Akt activation
Erratum in
-
Erratum to: Aryl hydrocarbon receptor/cytochrome P450 1A1 pathway mediates breast cancer stem cells expansion through PTEN inhibition and β-Catenin and Akt activation.Mol Cancer. 2017 Mar 24;16(1):69. doi: 10.1186/s12943-017-0636-5. Mol Cancer. 2017. PMID: 28340593 Free PMC article. No abstract available.
Abstract
Background: Breast cancer stem cells (CSCs) are small sub-type of the whole cancer cells that drive tumor initiation, progression and metastasis. Recent studies have demonstrated a role for the aryl hydrocarbon receptor (AhR)/cytochrome P4501A1 pathway in CSCs expansion. However, the exact molecular mechanisms remain unclear.
Methods: The current study was designed to a) determine the effect of AhR activation and inhibition on breast CSCs development, maintenance, self-renewal, and chemoresistance at the in vitro and in vivo levels and b) explore the role of β-Catenin, PI3K/Akt, and PTEN signaling pathways. To test this hypothesis, CSC characteristics of five human breast cancer cells; SKBR-3, MCF-7, and MDA-MB231, HS587T, and T47D treated with AhR activators or inhibitor were determined using Aldefluor assay, side population, and mammosphere formation. The mRNA, protein expression, cellular content and localization of the target genes were determined by RT-PCR, Western blot analysis, and Immunofluorescence, respectively. At the in vivo level, female Balb/c mice were treated with AhR/CYP1A1 inducer and histopathology changes and Immunohistochemistry examination for target proteins were determined.
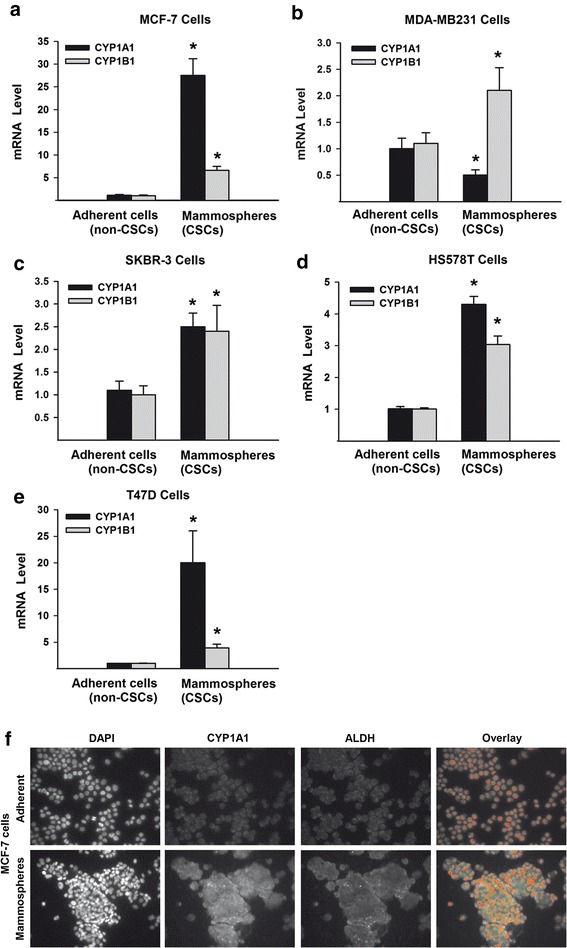
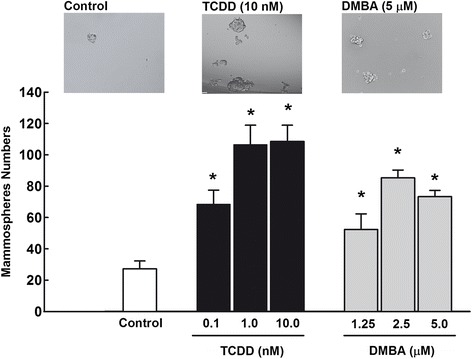
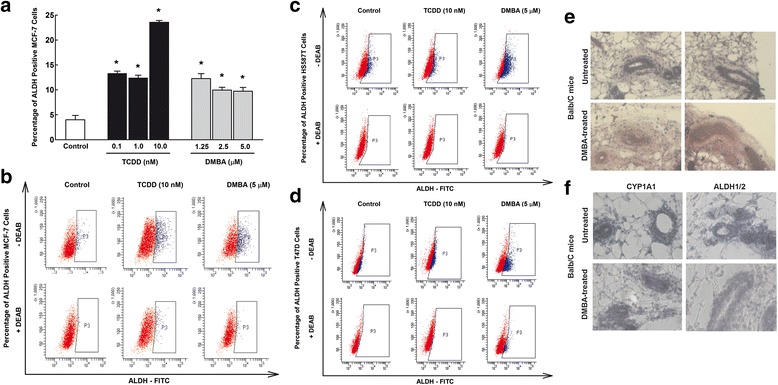
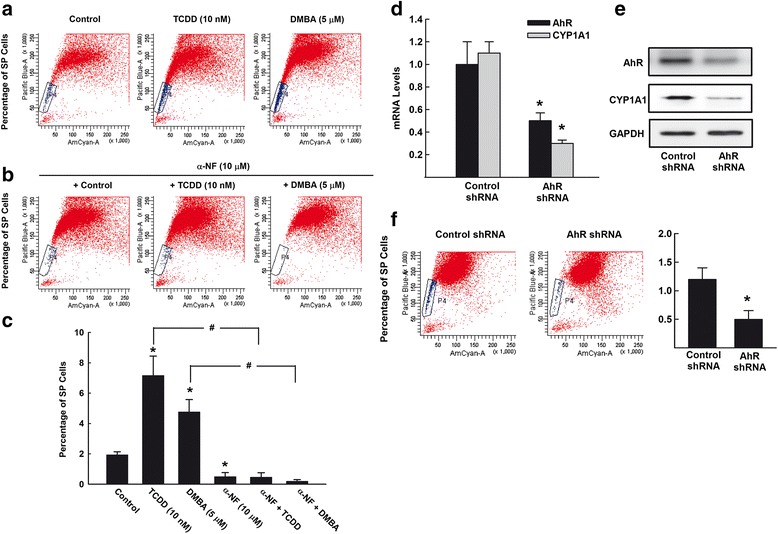
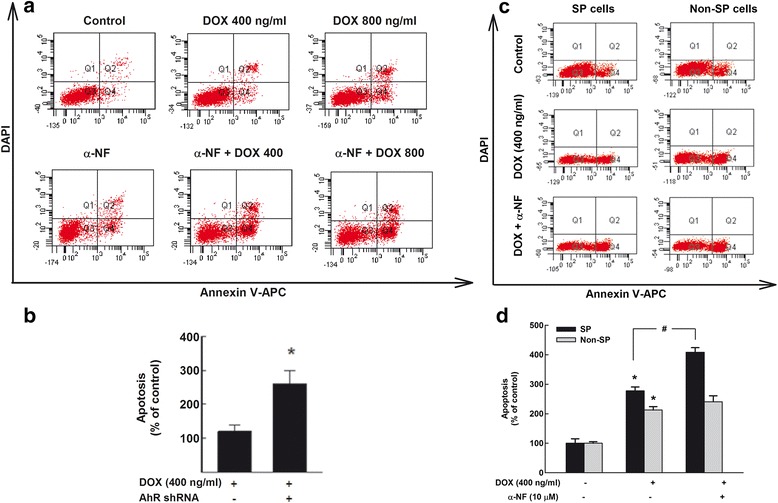
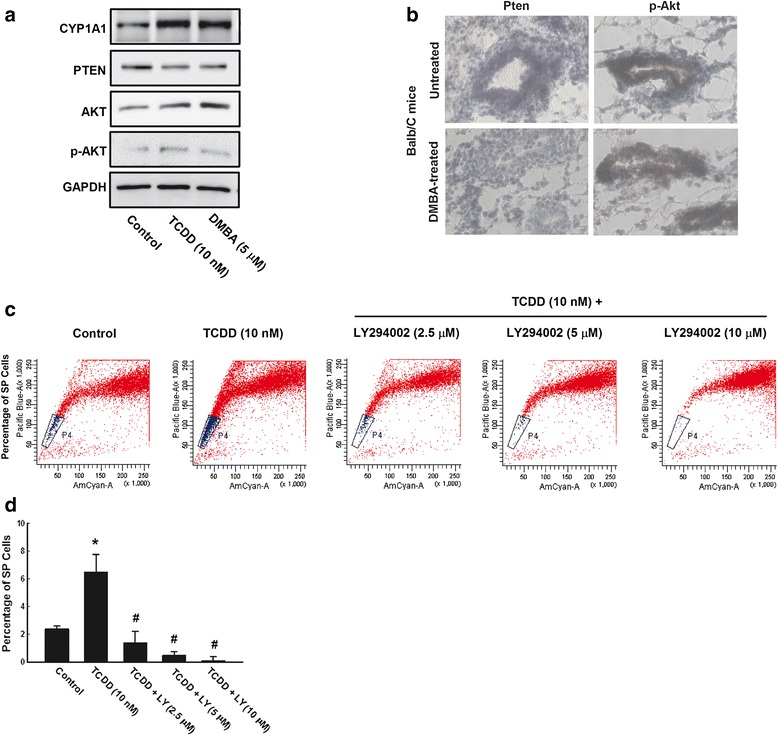
Results: The constitutive mRNA expression and cellular content of CYP1A1 and CYP1B1, AhR-regulated genes, were markedly higher in CSCs more than differentiating non-CSCs of five different human breast cancer cells. Activation of AhR/CYP1A1 in MCF-7 cells by TCDD and DMBA, strong AhR activators, significantly increased CSC-specific markers, mammosphere formation, aldehyde dehydrogenase (ALDH) activity, and percentage of side population (SP) cells, whereas inactivation of AhR/CYP1A1 using chemical inhibitor, α-naphthoflavone (α-NF), or by genetic shRNA knockdown, significantly inhibited the upregulation of ALDH activity and SP cells. Importantly, inactivation of the AhR/CYP1A1 significantly increased sensitization of CSCs to the chemotherapeutic agent doxorubicin. Mechanistically, Induction of AhR/CYP1A1 by TCDD and DMBA was associated with significant increase in β-Catenin mRNA and protein expression, nuclear translocation and its downstream target Cyclin D1, whereas AhR or CYP1A1 knockdown using shRNA dramatically inhibited β-Catenin cellular content and nuclear translocation. This was associated with significant inhibition of PTEN and induction of total and phosphorylated Akt protein expressions. Importantly, inhibition of PI3K/Akt pathway by LY294002 completely blocked the TCDD-induced SP cells expansion. In vivo, IHC staining of mammary gland structures of untreated and DMBA (30 mg/kg, IP)- treated mice, showed tremendous inhibition of PTEN expression accompanied with an increase in the expression p-Akt, β-Catenin and stem cells marker ALDH1.
Conclusions: The present study provides the first evidence that AhR/CYP1A1 signaling pathway is controlling breast CSCs proliferation, development, self-renewal and chemoresistance through inhibition of the PTEN and activation of β-Catenin and Akt pathways.
Keywords: AhR; Balb/c mice; Breast cancer; CYP1A1; Cancer stem cells; PI3K/Akt; PTEN; TCDD; shRNA; β-Catenin.
Figures



References
-
- Singh SK, Clarke ID, Terasaki M, Bonn VE, Hawkins C, Squire J, Dirks PB. Identification of a cancer stem cell in human brain tumors. Cancer Res. 2003;63:5821–8. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
