

Nuclear Dynamics of Heterochromatin Repair
- PMID: 28104289
- PMCID: PMC5285325
- DOI: 10.1016/j.tig.2016.12.004
Nuclear Dynamics of Heterochromatin Repair
Abstract
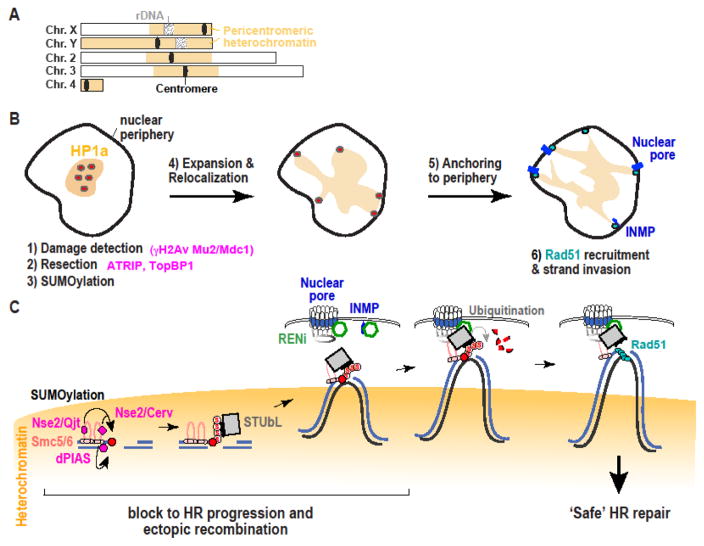
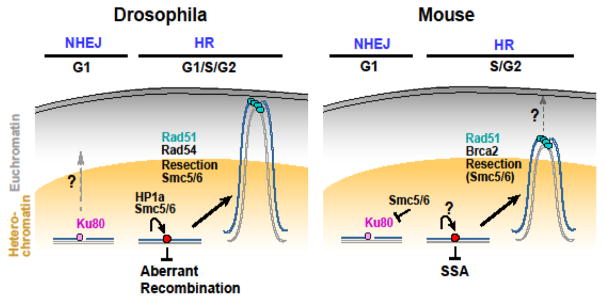
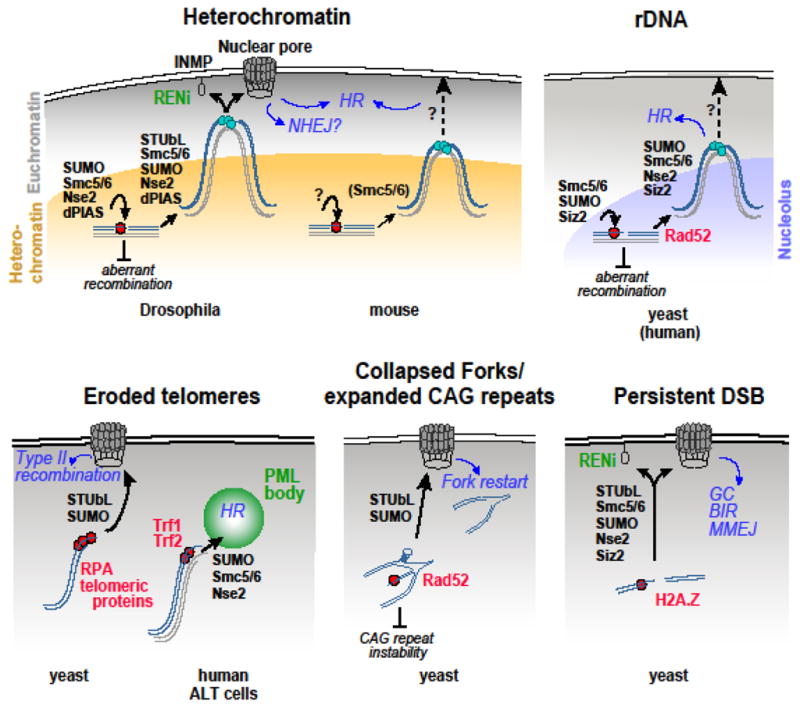
Repairing double-strand breaks (DSBs) is particularly challenging in pericentromeric heterochromatin, where the abundance of repeated sequences exacerbates the risk of ectopic recombination and chromosome rearrangements. Recent studies in Drosophila cells revealed that faithful homologous recombination (HR) repair of heterochromatic DSBs relies on the relocalization of DSBs to the nuclear periphery before Rad51 recruitment. We summarize here the exciting progress in understanding this pathway, including conserved responses in mammalian cells and surprising similarities with mechanisms in yeast that deal with DSBs in distinct sites that are difficult to repair, including other repeated sequences. We will also point out some of the most important open questions in the field and emerging evidence suggesting that deregulating these pathways might have dramatic consequences for human health.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Ho JW, Jung YL, Liu T, Alver BH, Lee S, Ikegami K, Sohn KA, Minoda A, Tolstorukov MY, Appert A, Parker SC, Gu T, Kundaje A, Riddle NC, Bishop E, Egelhofer TA, Hu SS, Alekseyenko AA, Rechtsteiner A, Asker D, Belsky JA, Bowman SK, Chen QB, Chen RA, Day DS, Dong Y, Dose AC, Duan X, Epstein CB, Ercan S, Feingold EA, Ferrari F, Garrigues JM, Gehlenborg N, Good PJ, Haseley P, He D, Herrmann M, Hoffman MM, Jeffers TE, Kharchenko PV, Kolasinska-Zwierz P, Kotwaliwale CV, Kumar N, Langley SA, Larschan EN, Latorre I, Libbrecht MW, Lin X, Park R, Pazin MJ, Pham HN, Plachetka A, Qin B, Schwartz YB, Shoresh N, Stempor P, Vielle A, Wang C, Whittle CM, Xue H, Kingston RE, Kim JH, Bernstein BE, Dernburg AF, Pirrotta V, Kuroda MI, Noble WS, Tullius TD, Kellis M, MacAlpine DM, Strome S, Elgin SC, Liu XS, Lieb JD, Ahringer J, Karpen GH, Park PJ. Comparative analysis of metazoan chromatin organization. Nature. 2014;512(7515):449–52. - PMC - PubMed
-
- Hoskins RA, Carlson JW, Wan KH, Park S, Mendez I, Galle SE, Booth BW, Pfeiffer BD, George RA, Svirskas R, Krzywinski M, Schein J, Accardo MC, Damia E, Messina G, Mendez-Lago M, de Pablos B, Demakova OV, Andreyeva EN, Boldyreva LV, Marra M, Carvalho AB, Dimitri P, Villasante A, Zhimulev IF, Rubin GM, Karpen GH, Celniker SE. The Release 6 reference sequence of the Drosophila melanogaster genome. Genome Res. 2015;25(3):445–58. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
