

A novel role for endothelial tetrahydrobiopterin in mitochondrial redox balance
- PMID: 28104455
- PMCID: PMC5338462
- DOI: 10.1016/j.freeradbiomed.2017.01.012
A novel role for endothelial tetrahydrobiopterin in mitochondrial redox balance
Abstract
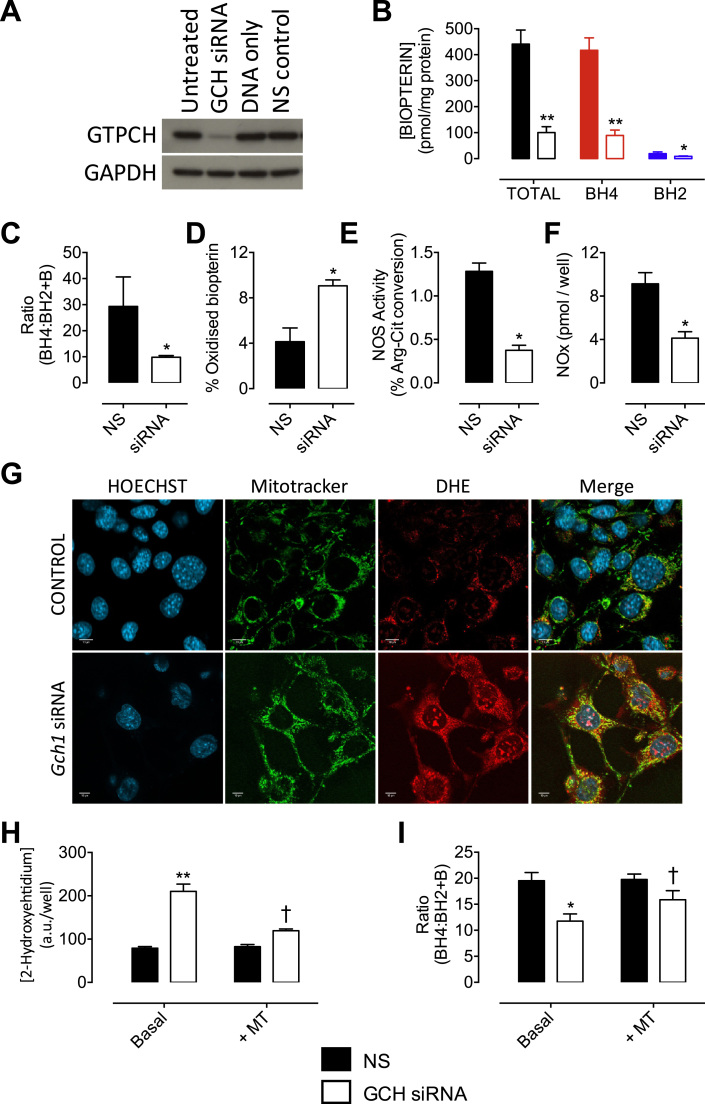
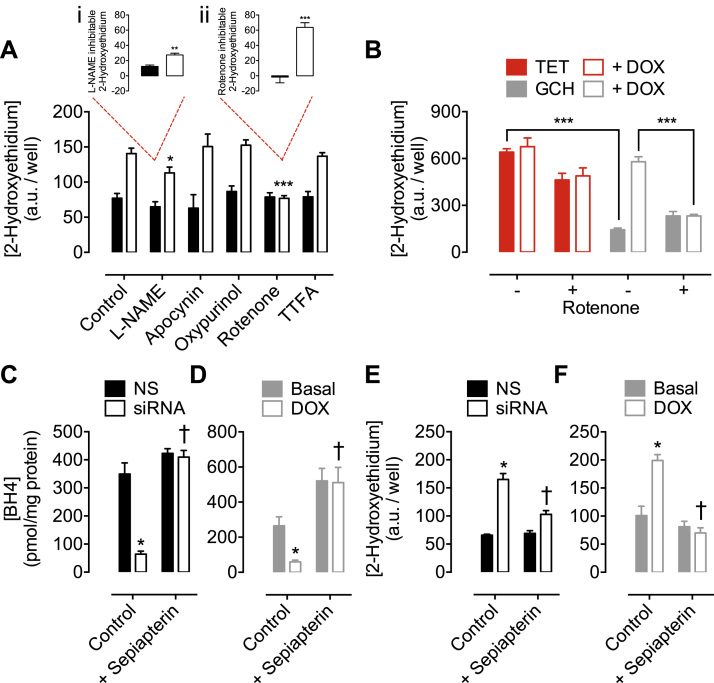
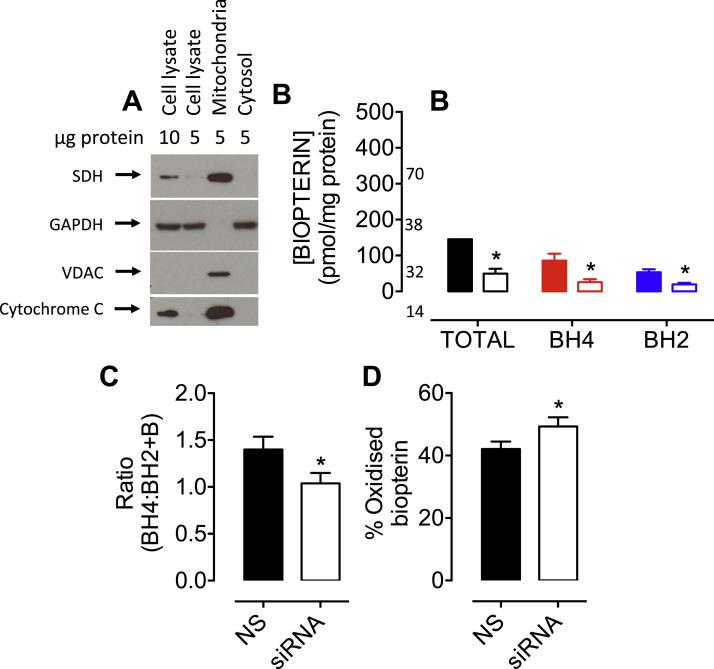
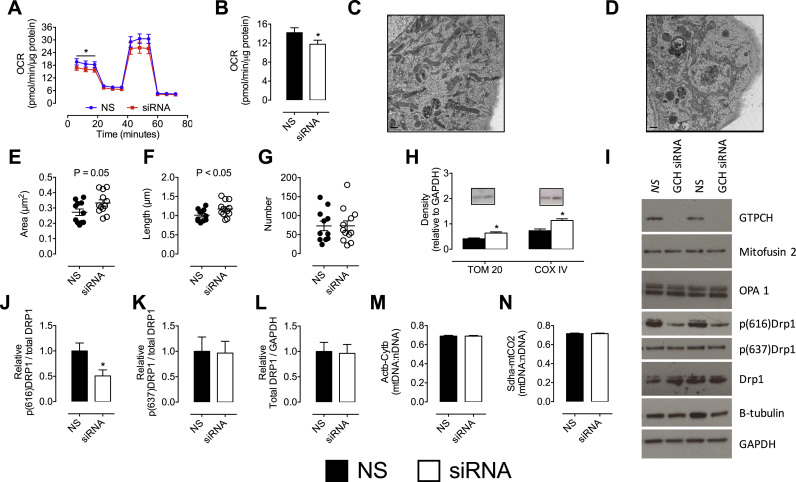
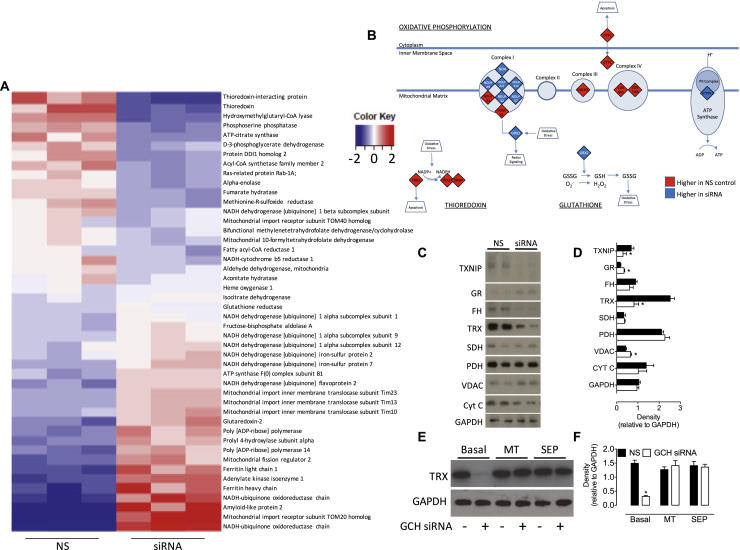
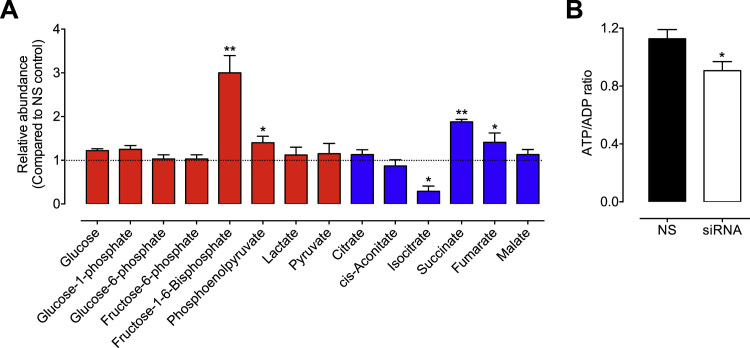
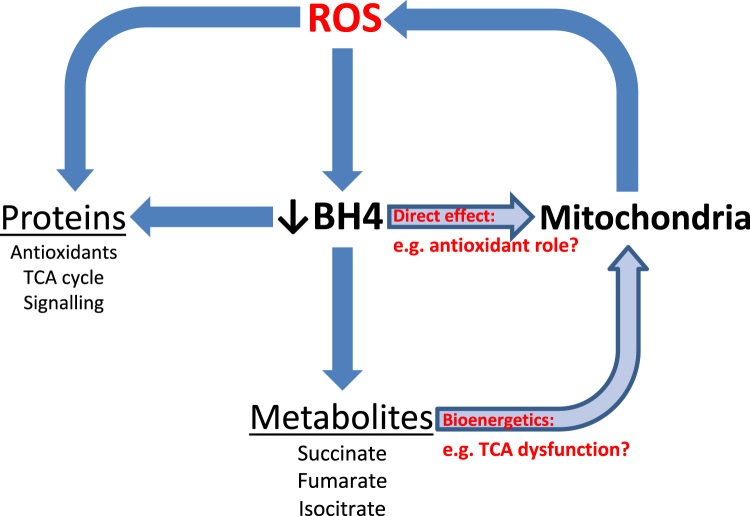
The redox co-factor tetrahydrobiopterin (BH4) regulates nitric oxide (NO) and reactive oxygen species (ROS) production by endothelial NOS (eNOS) and is an important redox-dependent signalling molecule in the endothelium. Loss of endothelial BH4 is observed in cardiovascular disease (CVD) states and results in decreased NO and increased superoxide (O2-) generation via eNOS uncoupling. Genetic mouse models of augmented endothelial BH4 synthesis have shown proof of concept that endothelial BH4 can alter CVD pathogenesis. However, clinical trials of BH4 therapy in vascular disease have been limited by systemic oxidation, highlighting the need to explore the wider roles of BH4 to find novel therapeutic targets. In this study, we aimed to elucidate the effects of BH4 deficiency on mitochondrial function and bioenergetics using targeted knockdown of the BH4 synthetic enzyme, GTP Cyclohydrolase I (GTPCH). Knockdown of GTPCH by >90% led to marked loss of cellular BH4 and a striking induction of O2- generation in the mitochondria of murine endothelial cells. This effect was likewise observed in BH4-depleted fibroblasts devoid of NOS, indicating a novel NOS-independent role for BH4 in mitochondrial redox signalling. Moreover, this BH4-dependent, mitochondria-derived ROS further oxidised mitochondrial BH4, concomitant with changes in the thioredoxin and glutathione antioxidant pathways. These changes were accompanied by a modest increase in mitochondrial size, mildly attenuated basal respiratory function, and marked changes in the mitochondrial proteome and cellular metabolome, including the accumulation of the TCA intermediate succinate. Taken together, these data reveal a novel NOS-independent role for BH4 in the regulation of mitochondrial redox signalling and bioenergetic metabolism.
Keywords: Mitochondria; Nitric oxide synthase; Redox state; Superoxide; Tetrahydrobiopterin.
Copyright © 2017. Published by Elsevier Inc.
Figures
References
-
- Kwon N.S., Nathan C.F., Stuehr D.J. Reduced biopterin as a cofactor in the generation of nitrogen oxides by murine macrophages. J. Biol. Chem. 1989;264:20496–20501. - PubMed
-
- Tayeh M.A., Marletta M.A. Macrophage oxidation of L-arginine to nitric oxide, nitrite, and nitrate. Tetrahydrobiopterin is required as a cofactor. J. Biol. Chem. 1989;264:19654–19658. - PubMed
-
- Stroes E., Hijmering M., van Zandvoort M., Wever R., Rabelink T.J., van Faassen E.E. Origin of superoxide production by endothelial nitric oxide synthase. FEBS Lett. 1998;438:161–164. - PubMed
-
- Kuzkaya N., Weissmann N., Harrison D.G., Dikalov S. Interactions of peroxynitrite, tetrahydrobiopterin, ascorbic acid, and thiols: implications for uncoupling endothelial nitric-oxide synthase. J. Biol. Chem. 2003;278:22546–22554. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
