

Lysosome-Dependent Activation of Human Dendritic Cells by the Vaccine Adjuvant QS-21
- PMID: 28105029
- PMCID: PMC5215313
- DOI: 10.3389/fimmu.2016.00663
Lysosome-Dependent Activation of Human Dendritic Cells by the Vaccine Adjuvant QS-21
Abstract
The adjuvant properties of the saponin QS-21 have been known for decades. It is a component of the Adjuvant System AS01 that is used in several vaccine candidates. QS-21 strongly potentiates both cellular and humoral immune responses to purified antigens, yet how it activates immune cells is largely unknown. Here, we report that QS-21 directly activated human monocyte-derived dendritic cells (moDCs) and promoted a pro-inflammatory transcriptional program. Cholesterol-dependent QS-21 endocytosis followed by lysosomal destabilization and Syk kinase activation were prerequisites for this response. Cathepsin B, a lysosomal cysteine protease, was essential for moDC activation in vitro and contributed to the adjuvant effects of QS-21 in vivo. Collectively, these findings provide new insights into the pathways involved in the direct activation of antigen-presenting cells by a clinically relevant QS-21 formulation.
Keywords: adjuvant; cathepsins; cytokine; dendritic cells; lysosome; saponins; vaccine.
Figures
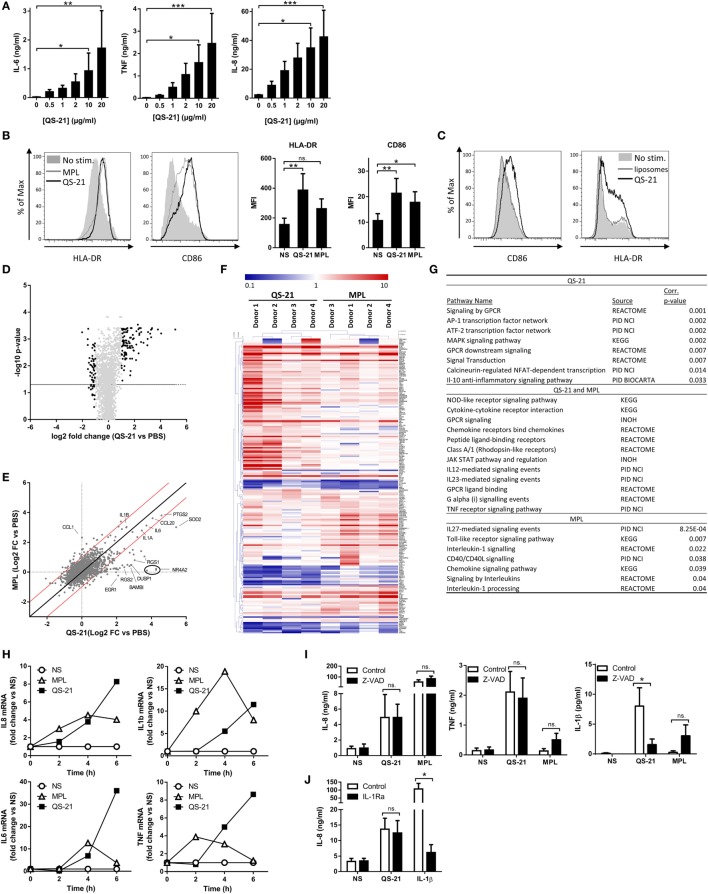
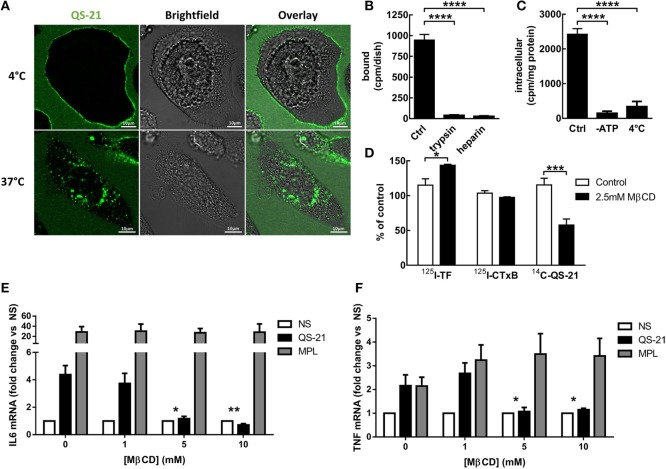
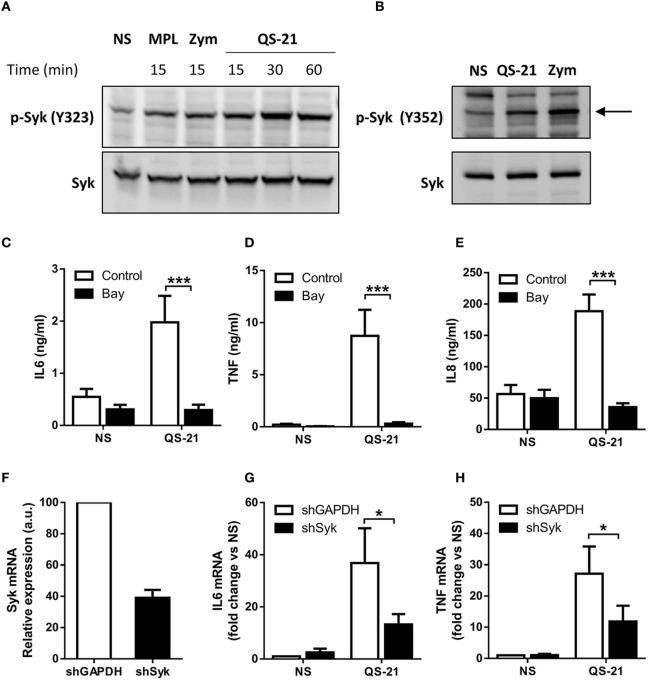
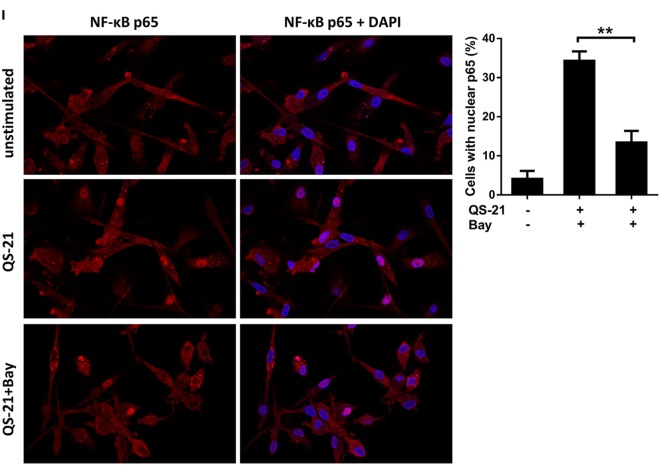
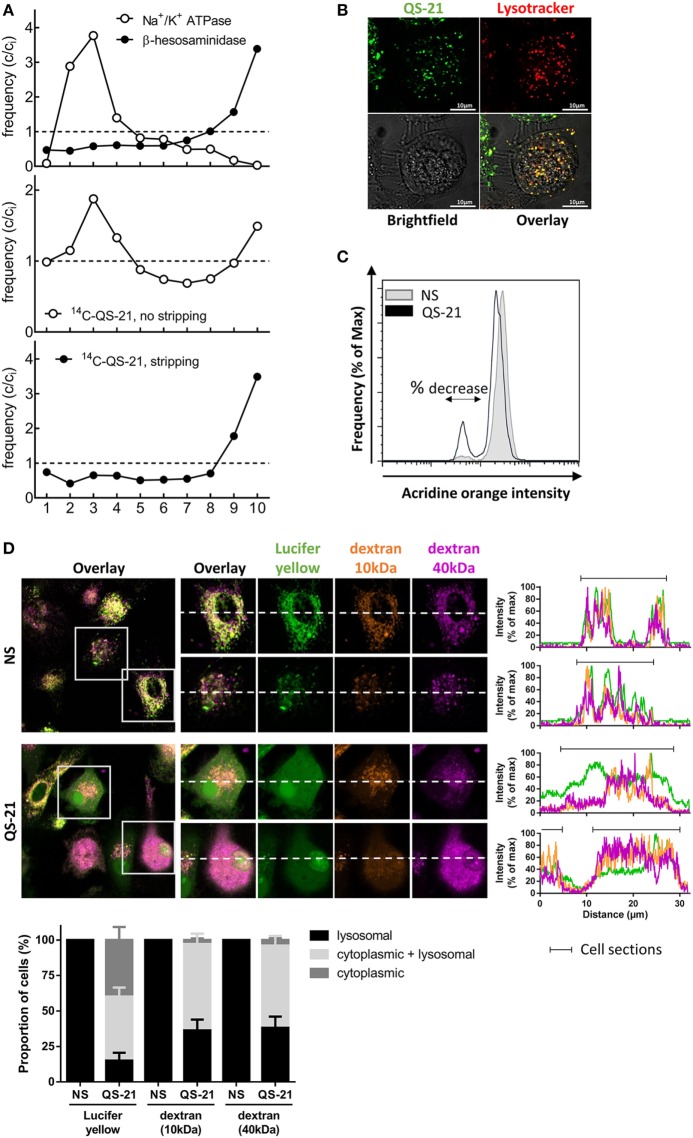
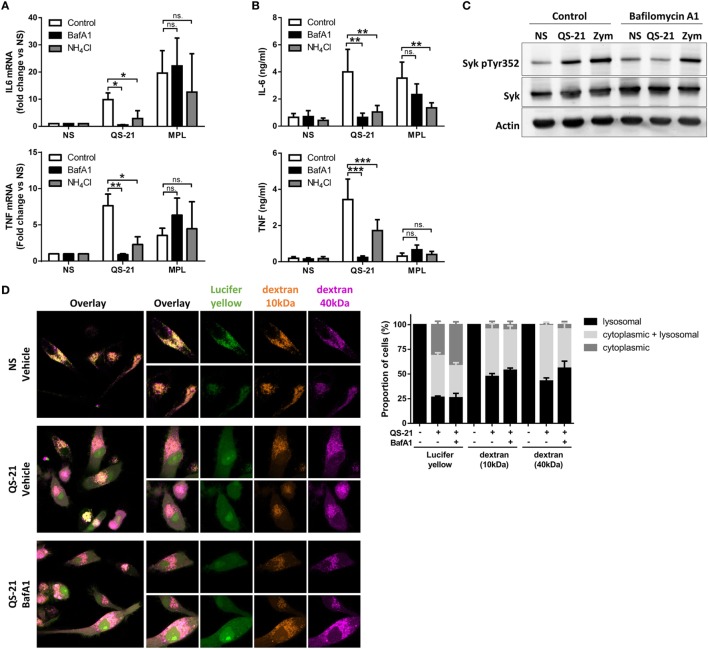
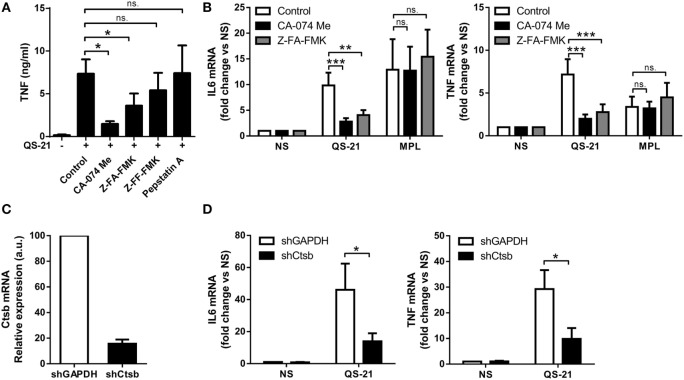
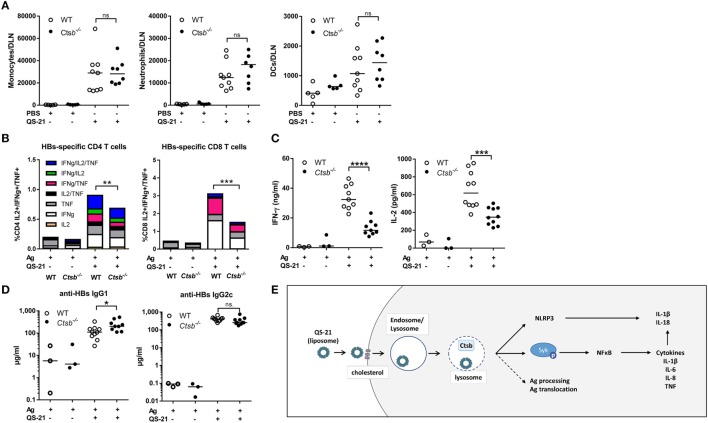
References
-
- Kensil CR, Patel U, Lennick M, Marciani D. Separation and characterization of saponins with adjuvant activity from Quillaja saponaria Molina cortex. J Immunol (1991) 146:431–7. - PubMed
-
- Kensil CR. Saponins as vaccine adjuvants. Crit Rev Ther Drug Carrier Syst (1996) 13:1–55. - PubMed
-
- Newman MJ, Wu JY, Gardner BH, Munroe KJ, Leombruno D, Recchia J, et al. Saponin adjuvant induction of ovalbumin-specific CD8+ cytotoxic T lymphocyte responses. J Immunol (1992) 148:2357–62. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous