

High-resolution cryo-EM structure of the proteasome in complex with ADP-AlFx
- PMID: 28106073
- PMCID: PMC5339833
- DOI: 10.1038/cr.2017.12
High-resolution cryo-EM structure of the proteasome in complex with ADP-AlFx
Abstract
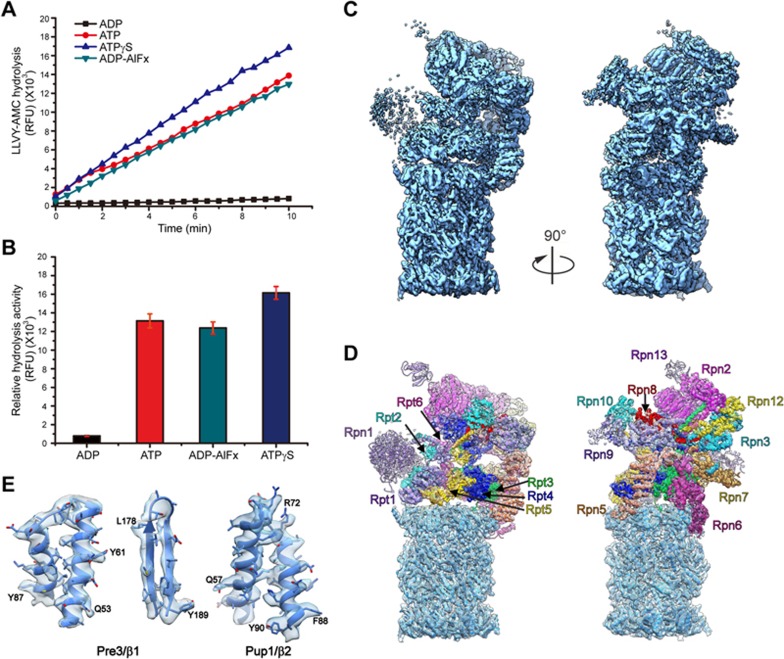
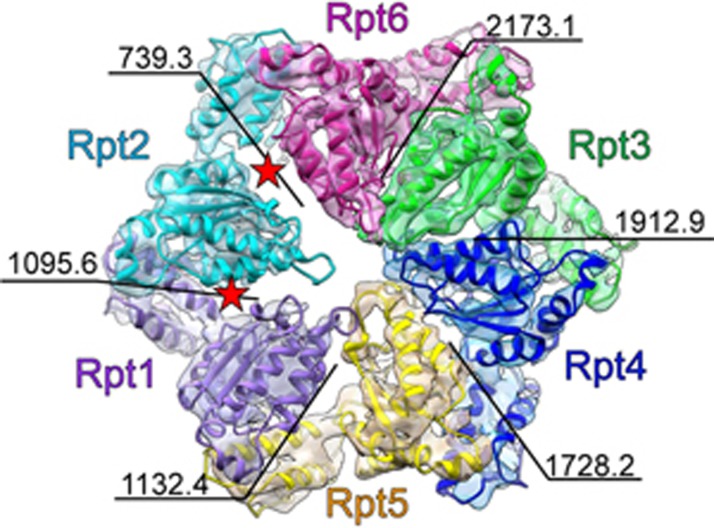
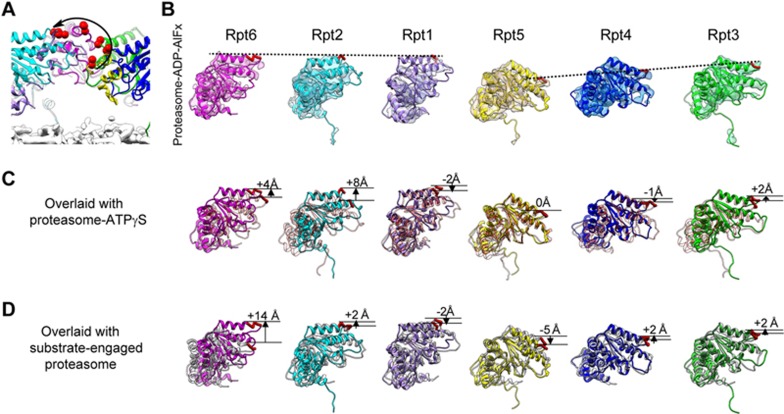
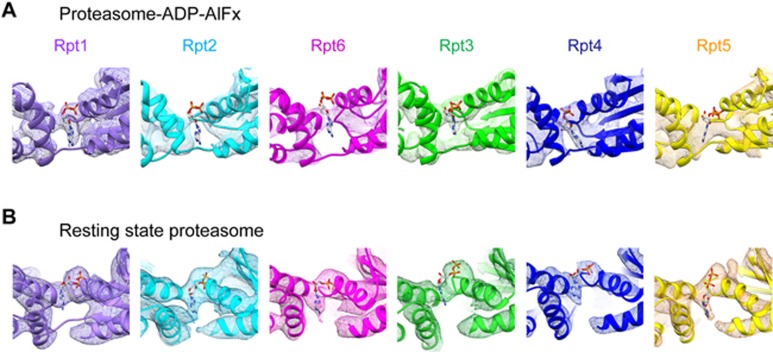
The 26S proteasome is an ATP-dependent dynamic 2.5 MDa protease that regulates numerous essential cellular functions through degradation of ubiquitinated substrates. Here we present a near-atomic-resolution cryo-EM map of the S. cerevisiae 26S proteasome in complex with ADP-AlFx. Our biochemical and structural data reveal that the proteasome-ADP-AlFx is in an activated state, displaying a distinct conformational configuration especially in the AAA-ATPase motor region. Noteworthy, this map demonstrates an asymmetric nucleotide binding pattern with four consecutive AAA-ATPase subunits bound with nucleotide. The remaining two subunits, Rpt2 and Rpt6, with empty or only partially occupied nucleotide pocket exhibit pronounced conformational changes in the AAA-ATPase ring, which may represent a collective result of allosteric cooperativity of all the AAA-ATPase subunits responding to ATP hydrolysis. This collective motion of Rpt2 and Rpt6 results in an elevation of their pore loops, which could play an important role in substrate processing of proteasome. Our data also imply that the nucleotide occupancy pattern could be related to the activation status of the complex. Moreover, the HbYX tail insertion may not be sufficient to maintain the gate opening of 20S core particle. Our results provide new insights into the mechanisms of nucleotide-driven allosteric cooperativity of the complex and of the substrate processing by the proteasome.
Figures
Comment in
-
Proteasomes, caught in the act.Cell Res. 2017 Mar;27(3):307-308. doi: 10.1038/cr.2017.22. Epub 2017 Feb 21. Cell Res. 2017. PMID: 28220774 Free PMC article.
Similar articles
-
Structure of an endogenous yeast 26S proteasome reveals two major conformational states.Proc Natl Acad Sci U S A. 2016 Mar 8;113(10):2642-7. doi: 10.1073/pnas.1601561113. Epub 2016 Feb 29. Proc Natl Acad Sci U S A. 2016. PMID: 26929360 Free PMC article.
-
Structure of the 26S proteasome with ATP-γS bound provides insights into the mechanism of nucleotide-dependent substrate translocation.Proc Natl Acad Sci U S A. 2013 Apr 30;110(18):7264-9. doi: 10.1073/pnas.1305782110. Epub 2013 Apr 15. Proc Natl Acad Sci U S A. 2013. PMID: 23589842 Free PMC article.
-
Structural mechanism for nucleotide-driven remodeling of the AAA-ATPase unfoldase in the activated human 26S proteasome.Nat Commun. 2018 Apr 10;9(1):1360. doi: 10.1038/s41467-018-03785-w. Nat Commun. 2018. PMID: 29636472 Free PMC article.
-
Recent advances in the structural biology of the 26S proteasome.Int J Biochem Cell Biol. 2016 Oct;79:437-442. doi: 10.1016/j.biocel.2016.08.008. Epub 2016 Aug 4. Int J Biochem Cell Biol. 2016. PMID: 27498189 Review.
-
Structure, Dynamics and Function of the 26S Proteasome.Subcell Biochem. 2021;96:1-151. doi: 10.1007/978-3-030-58971-4_1. Subcell Biochem. 2021. PMID: 33252727 Review.
Cited by
-
Structure and Function of the 26S Proteasome.Annu Rev Biochem. 2018 Jun 20;87:697-724. doi: 10.1146/annurev-biochem-062917-011931. Epub 2018 Apr 13. Annu Rev Biochem. 2018. PMID: 29652515 Free PMC article. Review.
-
The conformational landscape of TRiC ring-opening and its underlying stepwise mechanism revealed by cryo-EM.QRB Discov. 2024 Dec 16;6:e7. doi: 10.1017/qrd.2024.17. eCollection 2025. QRB Discov. 2024. PMID: 40070846 Free PMC article.
-
Mechanisms and regulation of substrate degradation by the 26S proteasome.Nat Rev Mol Cell Biol. 2025 Feb;26(2):104-122. doi: 10.1038/s41580-024-00778-0. Epub 2024 Oct 3. Nat Rev Mol Cell Biol. 2025. PMID: 39362999 Review.
-
ISG15 suppresses ovulation and female fertility by ISGylating ADAMTS1.Cell Biosci. 2023 May 11;13(1):84. doi: 10.1186/s13578-023-01024-4. Cell Biosci. 2023. PMID: 37170317 Free PMC article.
-
Cryo-EM Reveals Unanchored M1-Ubiquitin Chain Binding at hRpn11 of the 26S Proteasome.Structure. 2020 Nov 3;28(11):1206-1217.e4. doi: 10.1016/j.str.2020.07.011. Epub 2020 Aug 11. Structure. 2020. PMID: 32783951 Free PMC article.
References
-
- Schwartz AL, Ciechanover A. Targeting proteins for destruction by the ubiquitin system: implications for human pathobiology. Annu Rev Pharmacol Toxicol 2009; 49:73–96. - PubMed
-
- Goldberg AL. Nobel committee tags ubiquitin for distinction. Neuron 2005; 45:339–344. - PubMed
-
- Glickman MH, Ciechanover A. The ubiquitin-proteasome proteolytic pathway: destruction for the sake of construction. Physiol Rev 2002; 82:373–428. - PubMed
-
- Leroy E, Boyer R, Auburger G, et al. The ubiquitin pathway in Parkinson's disease. Nature 1998; 395:451–452. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
