

Rapid generation of hypomorphic mutations
- PMID: 28106166
- PMCID: PMC5263891
- DOI: 10.1038/ncomms14112
Rapid generation of hypomorphic mutations
Erratum in
-
Corrigendum: Rapid generation of hypomorphic mutations.Nat Commun. 2017 Feb 16;8:14705. doi: 10.1038/ncomms14705. Nat Commun. 2017. PMID: 28205645 Free PMC article. No abstract available.
Abstract
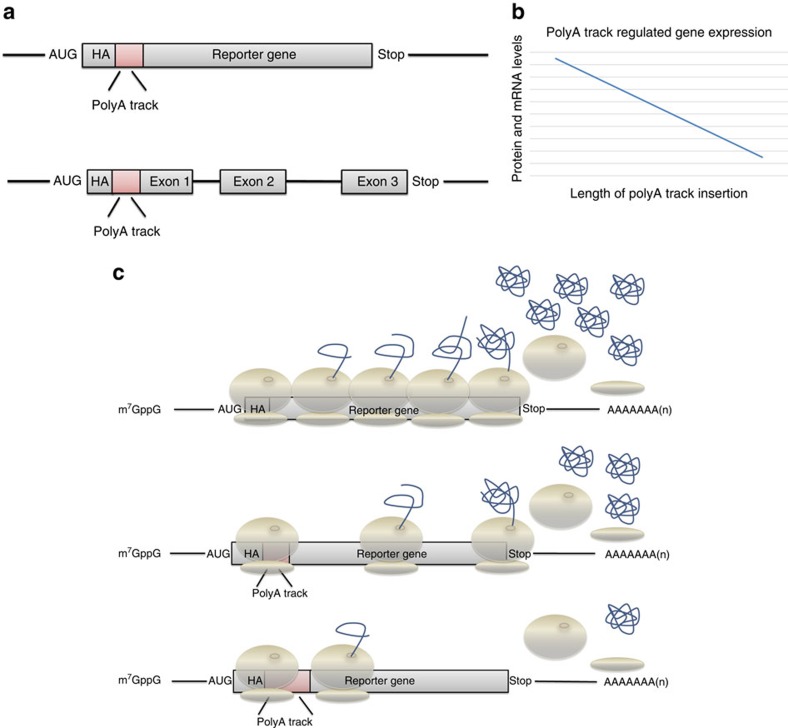
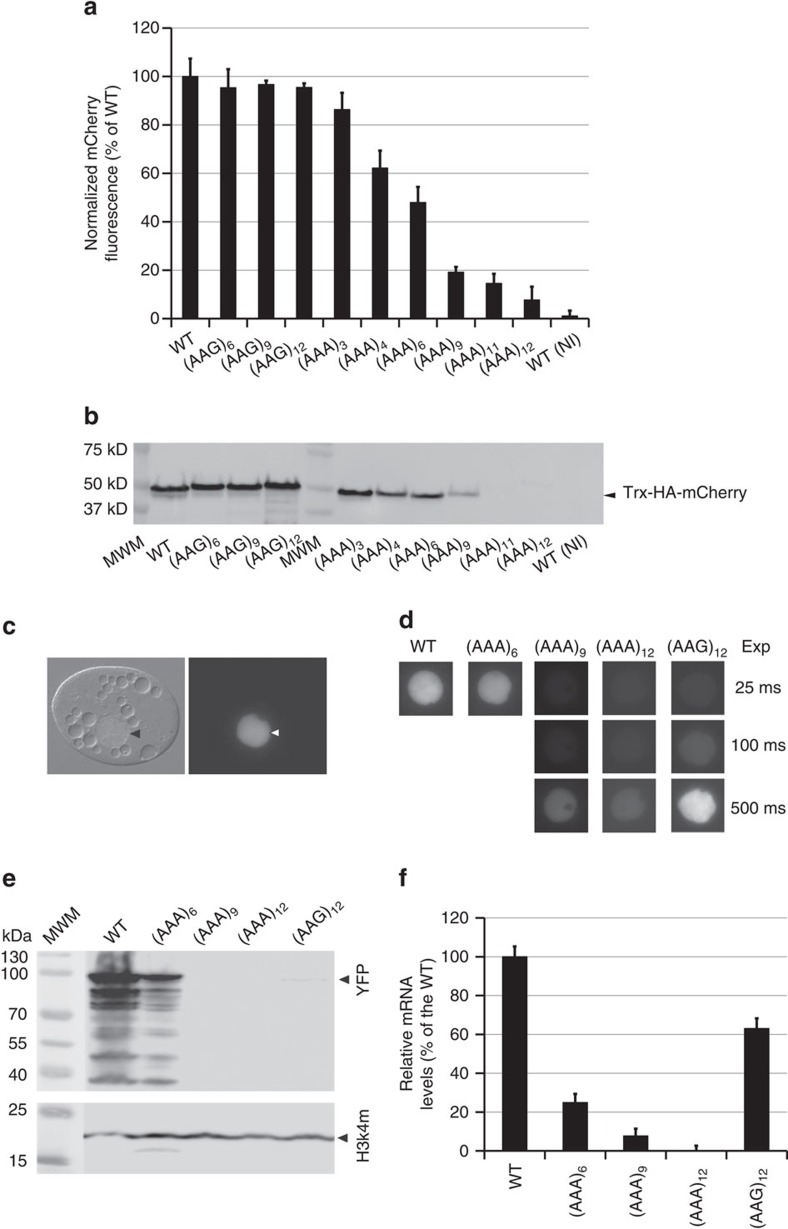
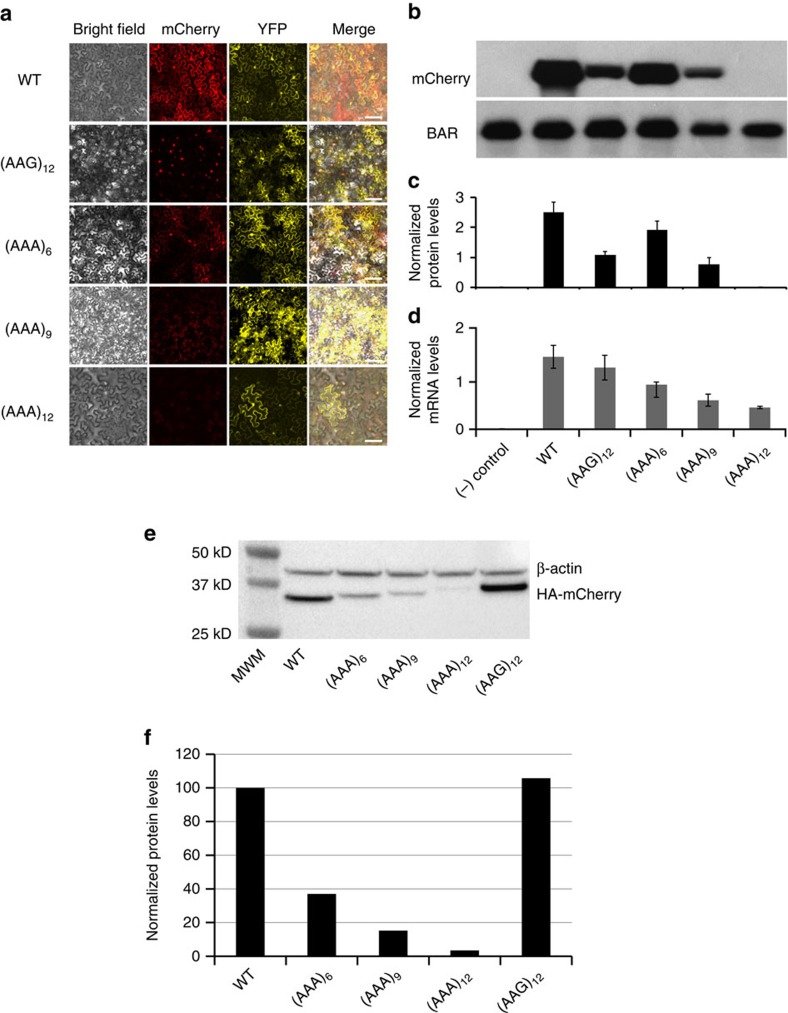
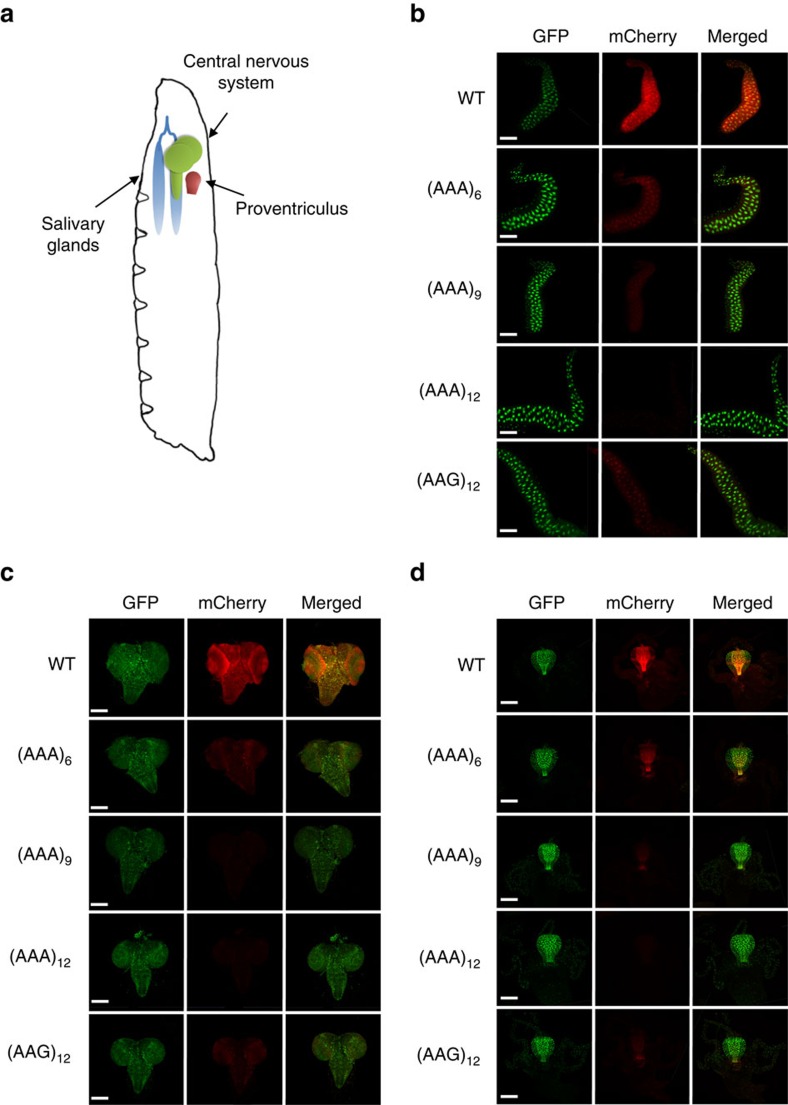
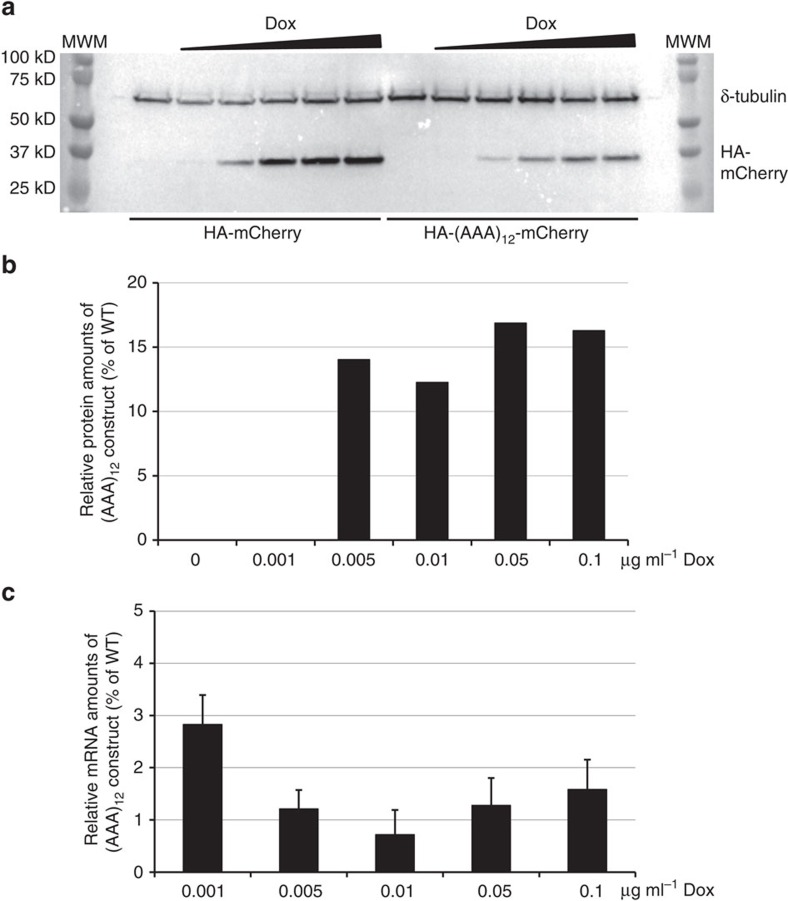
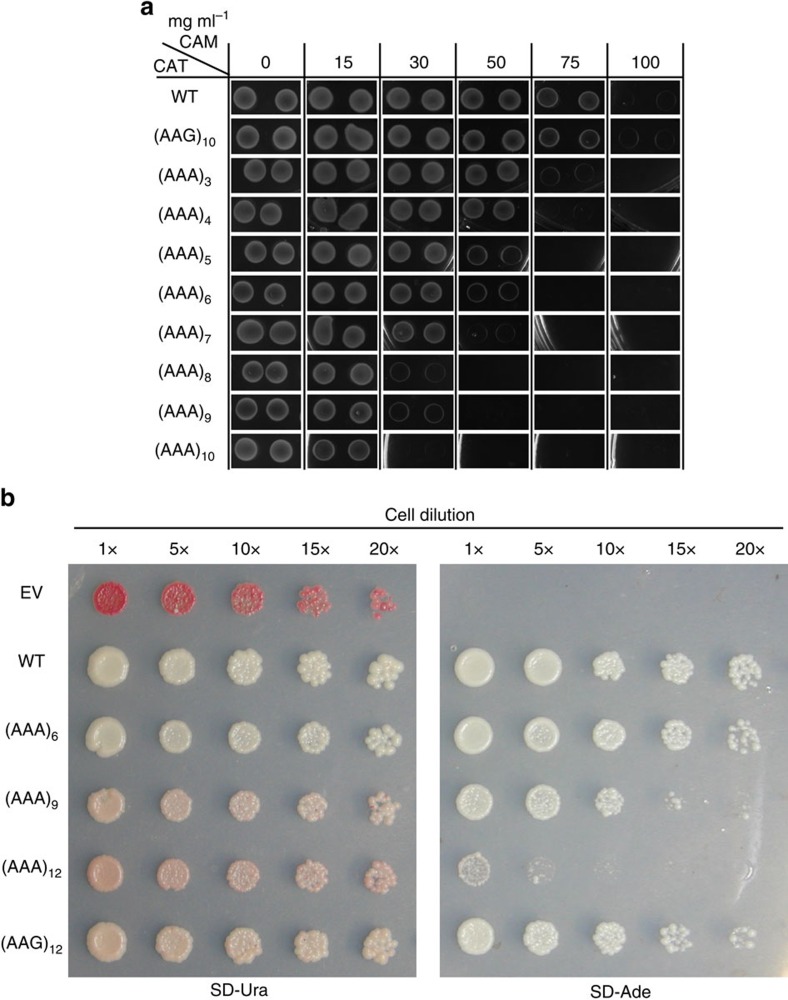
Hypomorphic mutations are a valuable tool for both genetic analysis of gene function and for synthetic biology applications. However, current methods to generate hypomorphic mutations are limited to a specific organism, change gene expression unpredictably, or depend on changes in spatial-temporal expression of the targeted gene. Here we present a simple and predictable method to generate hypomorphic mutations in model organisms by targeting translation elongation. Adding consecutive adenosine nucleotides, so-called polyA tracks, to the gene coding sequence of interest will decrease translation elongation efficiency, and in all tested cell cultures and model organisms, this decreases mRNA stability and protein expression. We show that protein expression is adjustable independent of promoter strength and can be further modulated by changing sequence features of the polyA tracks. These characteristics make this method highly predictable and tractable for generation of programmable allelic series with a range of expression levels.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Muller H. J. Further studies on the nature and causes of gene mutations. Proc. 6th Int. Congr. Genet. 1, 213–255 (1932).
-
- Bonde M. T. et al. Predictable tuning of protein expression in bacteria. Nat. Methods 13, 233–236 (2016). - PubMed
-
- Chappell J., Watters K. E., Takahashi M. K. & Lucks J. B. A renaissance in RNA synthetic biology: New mechanisms, applications and tools for the future. Curr. Opin. Chem. Biol. 28, 47–56 (2015). - PubMed
-
- Dawlaty M. M. & van Deursen J. M. Gene targeting methods for studying nuclear transport factors in mice. Methods 39, 370–378 (2006). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
