

Review
doi: 10.1101/cshperspect.a022186.
TGF-β Family Signaling in Embryonic and Somatic Stem-Cell Renewal and Differentiation
Affiliations
- PMID: 28108485
- PMCID: PMC5495062
- DOI: 10.1101/cshperspect.a022186
Item in Clipboard
Review
TGF-β Family Signaling in Embryonic and Somatic Stem-Cell Renewal and Differentiation
Cold Spring Harb Perspect Biol.
.
Abstract
Soon after the discovery of transforming growth factor-β (TGF-β), seminal work in vertebrate and invertebrate models revealed the TGF-β family to be central regulators of tissue morphogenesis. Members of the TGF-β family direct some of the earliest cell-fate decisions in animal development, coordinate complex organogenesis, and contribute to tissue homeostasis in the adult. Here, we focus on the role of the TGF-β family in mammalian stem-cell biology and discuss its wide and varied activities both in the regulation of pluripotency and in cell-fate commitment.
Copyright © 2017 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
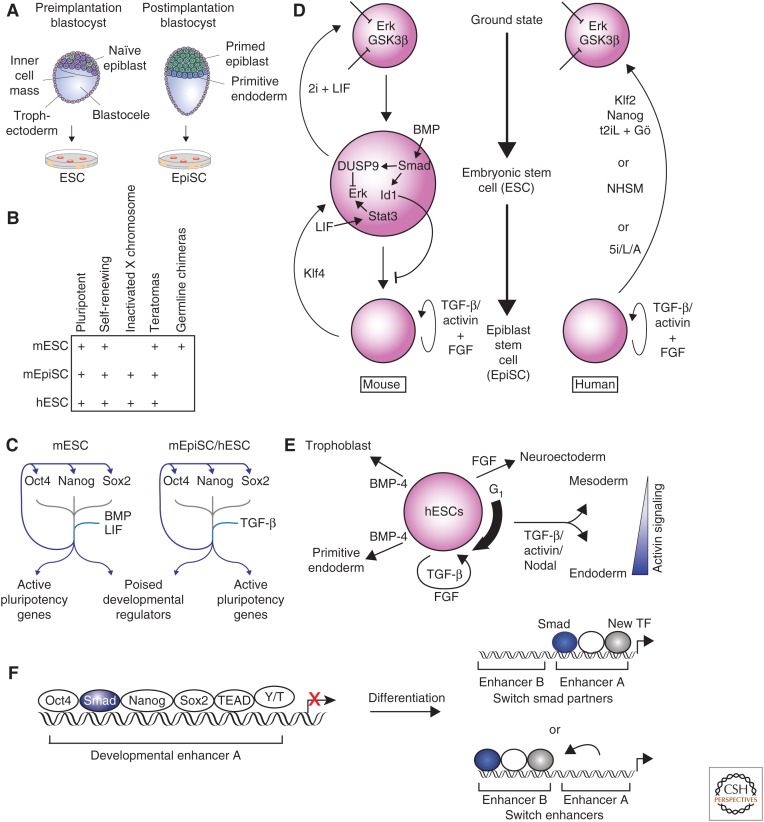
Transforming growth factor β (TGF-β) family signaling in embryonic stem cells (ESCs). (A) Mouse embryonic stem cells (mESCs) are derived from naïve epiblast cells in the preimplantation blastocyst, whereas postimplantation epiblasts (EpiSCs) are derived from primed epiblast cells in the postimplantation blastocyst. (B) The defining characteristics of mESCs, mEpiSCs, and human (h) ESCs are shown. (C) Oct4 (Pou5f1), Nanog, and Sox2 are the key transcription factors that maintain ESC state. These factors co-occupy the genome at their own promoters, at the promoters of other key ESC genes, and at developmental regulators that are repressed but poised to be activated during differentiation. Bone morphogenetic protein (BMP) and leukemia inhibitory factor (LIF) reinforce this network in mESCs, and TGF-β family signaling reinforces this network in hESCs and EpiSCs. (D) Key pathways that interact with TGF-β family signaling to maintain mESC (left) and hESC (right) states are shown. BMP signaling is a key factor in the maintenance of mESC state, whereas TGF-β and/or activin maintain the mEpiSC and hESC states. (E) The role of TGF-β family signaling in hESCs differentiation is indicated. The thick arrow to the right of the hESC indicates that hESCs respond most efficiently to activin signaling with Smad2 and Smad3 activation during early G1 phase of cell cycle. Once cells differentiate into mesendoderm, the concentration of activin is a major determining factor in the differentiation toward mesoderm or endoderm. (F) Smad2 and/or Smad3 co-occupy the genome with the key transcription factors that maintain the ESC state (left). During differentiation into endoderm, the expression of the key ESC transcription factors is lost. Smad2 and/or Smad3 regulate induction of new genes by either continuing to occupy the same enhancer but with different combinations of transcription factors (top right) or by moving to new enhancers in association with new transcription factors (bottom right). Y/T, YAP and TAZ.
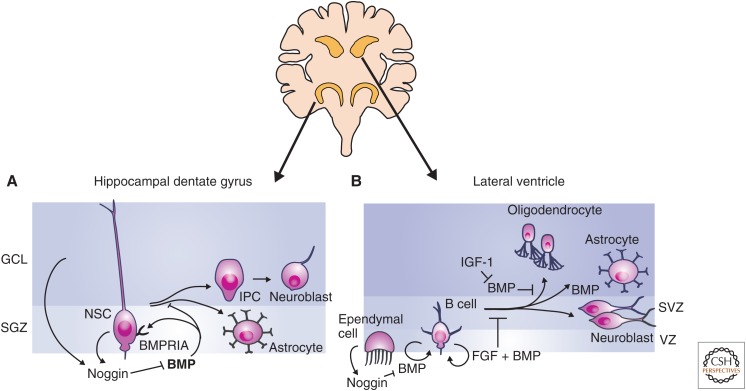
Transforming growth factor β (TGF-β) family signaling in neural stem cells. Adult neural stem cells (NSCs) are primarily located in the subgranular zone (SGZ) of the hippocampal dentate gyrus and the subventricular zone (SVZ) lining the lateral ventricles. (A) Bone morphogenetic protein (BMP) signaling promotes the maintenance of the NSC state in the SGZ, and signaling through BMPRIA inhibits NSC proliferation. NSCs of the SGZ produce the BMP inhibitor Noggin. Loss of BMP signaling through repression of BMPRIA or inhibition of BMP signaling results in the formation of intermediate progenitor cells (IPCs), which differentiate into neuroblasts and astrocytes in vivo. BMP is drawn in bold in the dentate gyrus to indicate increased levels of BMP expression relative to SVZ. (B) In the SVZ, NSCs are called B cells and are maintained as slowly cycling NSCs by BMP and fibroblast growth factor (FGF) signaling. Noggin is produced by ependymal cells and inhibits of BMP signaling. BMP signaling promotes maintenance of the NSC state, but once cells begin to proliferate and differentiate, BMP signaling can also promote astrocyte differentiation. GCL, Granular cell layer.
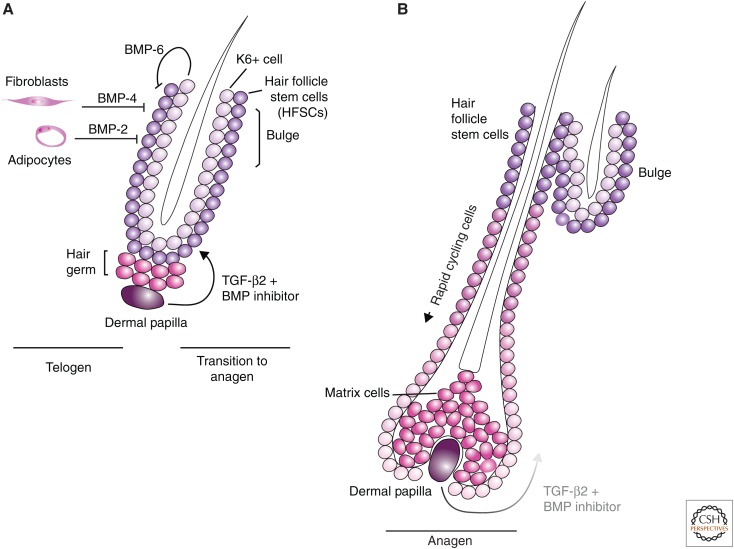
Transforming growth factor β (TGF-β) family signaling in hair follicle stem cells. (A) Telogen represents the quiescent state of hair follicle development. Hair follicle stem cells (HFSCs) are located in the bulge. Cells including dermal fibroblasts and adipocytes and K6+ cells secrete bone morphogenetic protein (BMP) molecules, which maintain HFSCs in their slowly proliferating state. Expression of Noggin and TGF-β from the dermal papilla (DP) cells blocks BMP signaling and promotes proliferation of the HFSCs. The progenitor cells that differentiate from the HFSCs are responsible for growth of the hair follicle. (B) During anagen, the hair follicle grows, pushing the DP and the TGF-β2 produced by these cells further from the HFSCs in the bulge. At a certain point, the BMP signaling wins out over the TGF-β signal, resulting in HFSC quiescence and atrophy of the hair follicle.
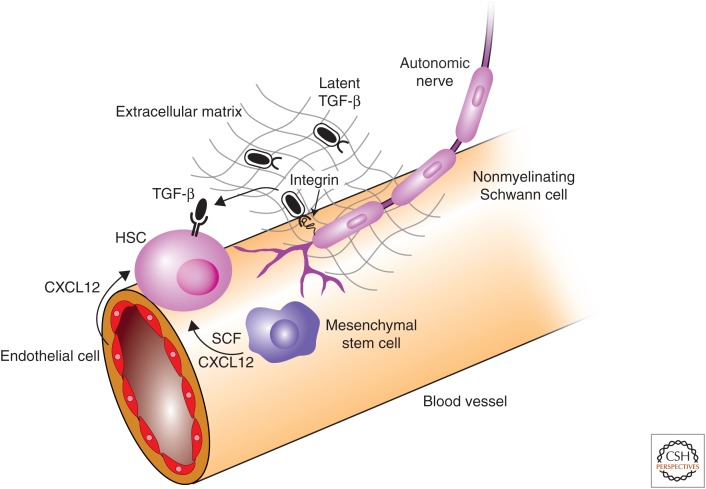
Transforming growth factor β (TGF-β) family signaling in hematopoietic stem cells. The niche provides signaling support to maintain hematopoietic stem-cell (HSC) state. CXCL12 and stem-cell factor (SCF) are both required for HSC maintenance and are provided by mesenchymal stem cells (MSCs) and endothelial cells. Inactive (latent) TGF-β is produce by HSCs and other cells in the niche. Nonmyelinating Schwann cells express integrin αvβ8, which binds the latent TGF-β complex and recruits metalloproteinases that cleave latent TGF-β and release active TGF-β. TGF-β acts through TβRII on the surface of HCSs to inhibit proliferation and promote maintenance of HSC state.
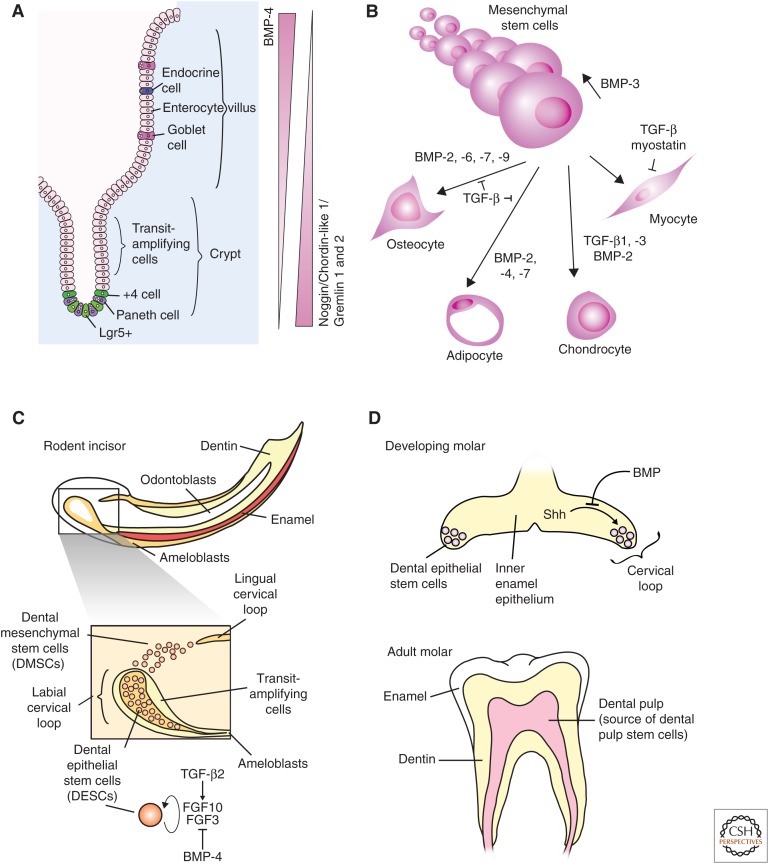
Transforming growth factor β (TGF-β) family signaling in intestinal, mesenchymal, and dental stem cells. (A) Bone morphogenic protein 4 (BMP-4) signaling promotes differentiation of intestinal stem cells, whereas inhibition of BMP signaling by Noggin, Gremlin 1 or 2, and Chordin-like 1 promote maintenance of the intestinal stem-cell fate. (B) Mesenchymal stem-cell (MSC) maintenance and differentiation is regulated by BMP and TGF-β signaling. BMP-3 signaling promotes MSC proliferation and the indicated BMPs promote osteocyte and adipocyte differentiation. TGF-β1 and -β3 and BMP-2 coordinate to promote chondrocyte differentiation, and both myostatin and TGF-β inhibit myoblast differentiation. (C) Rodent incisors continue to grow throughout life, and growth is maintained by dental epithelial stem cells (DESCs) and dental mesenchymal stem cells (DMSCs). DESCs differentiate into ameloblasts to produce enamel, and DMSCs differentiate into odontoblasts to produce dentin. TGF-β2 increases and BMP-4 represses expression of fibroblast growth factor 3 (FGF3) and FGF10. FGF3 and FGF10 inhibit differentiation and promote proliferation of DESCs. (D) Dental stem cells are present in the developing molar and are lost in the adult molar. Shh promotes maintenance of the DESCs in the developing molar until the signal is blocked by BMP (top). The adult molar does not contain DESCs, but the dental pulp is a source of MSCs (bottom).
References
-
- Ahrens M, Ankenbauer T, Schröder D, Hollnagel A, Mayer H, Gross G. 1993. Expression of human bone morphogenetic proteins-2 or -4 in murine mesenchymal progenitor C3H10T1/2 cells induces differentiation into distinct mesenchymal cell lineages. DNA Cell Biol 12: 871–880. - PubMed
-
- Aizawa K, Ageyama N, Terao K, Hisatsune T. 2011. Primate-specific alterations in neural stem/progenitor cells in the aged hippocampus. Neurobiol Aging 32: 140–150. - PubMed
-
- Amit M, Carpenter MK, Inokuma MS, Chiu CP, Harris CP, Waknitz MA, Itskovitz-Eldor J, Thomson JA. 2000. Clonally derived human embryonic stem cell lines maintain pluripotency and proliferative potential for prolonged periods of culture. Dev Biol 227: 271–278. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources