

mGluR2 versus mGluR3 Metabotropic Glutamate Receptors in Primate Dorsolateral Prefrontal Cortex: Postsynaptic mGluR3 Strengthen Working Memory Networks
- PMID: 28108498
- PMCID: PMC5974790
- DOI: 10.1093/cercor/bhx005
mGluR2 versus mGluR3 Metabotropic Glutamate Receptors in Primate Dorsolateral Prefrontal Cortex: Postsynaptic mGluR3 Strengthen Working Memory Networks
Abstract
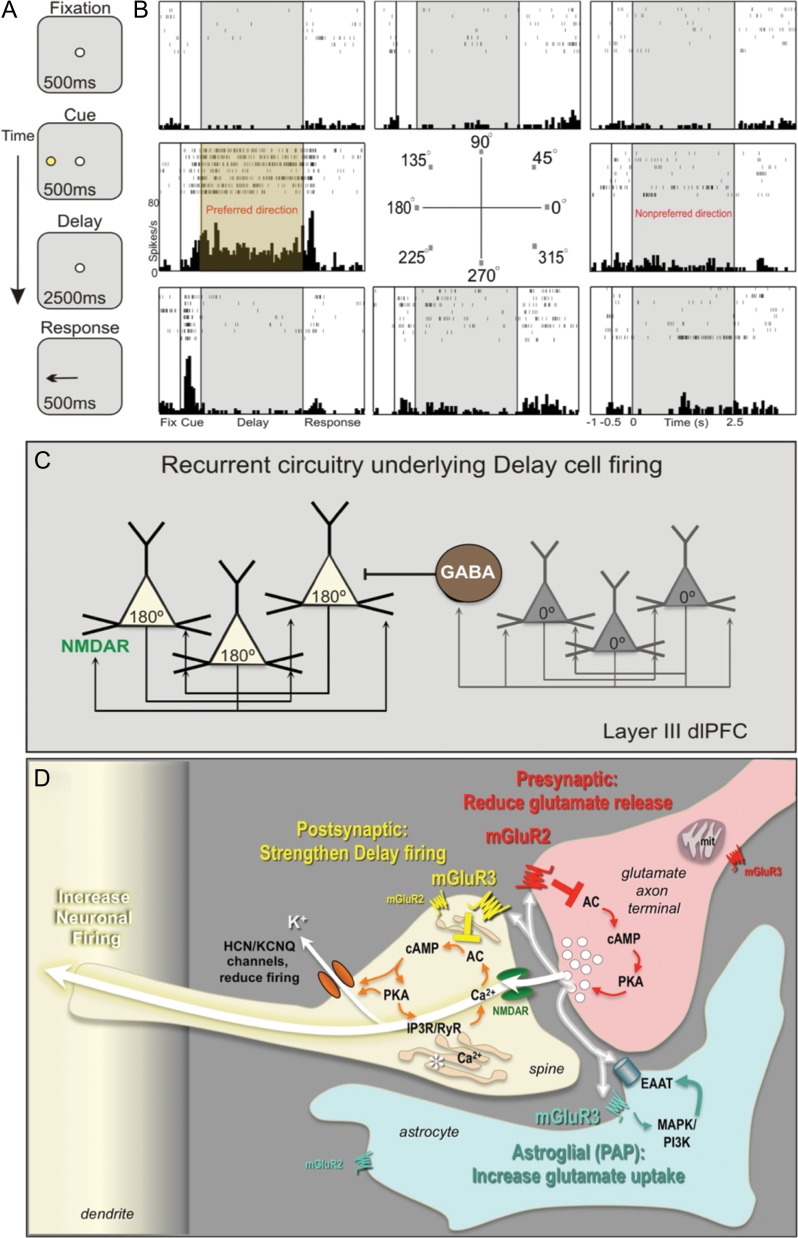
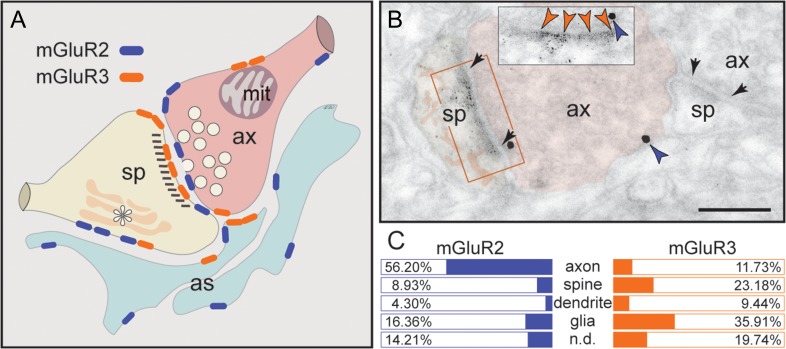
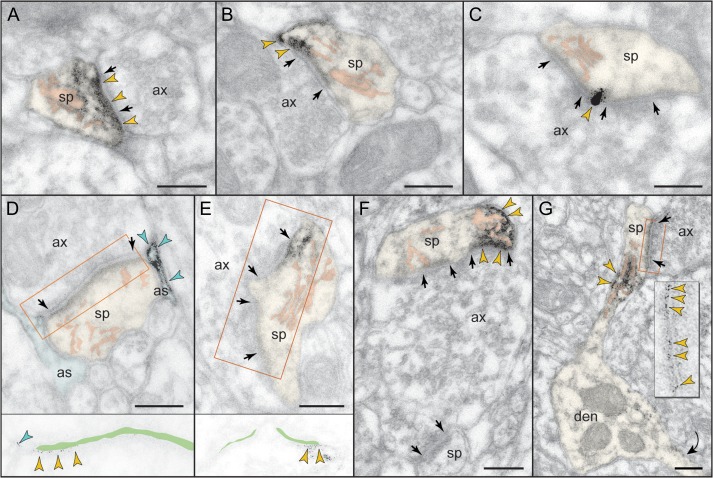
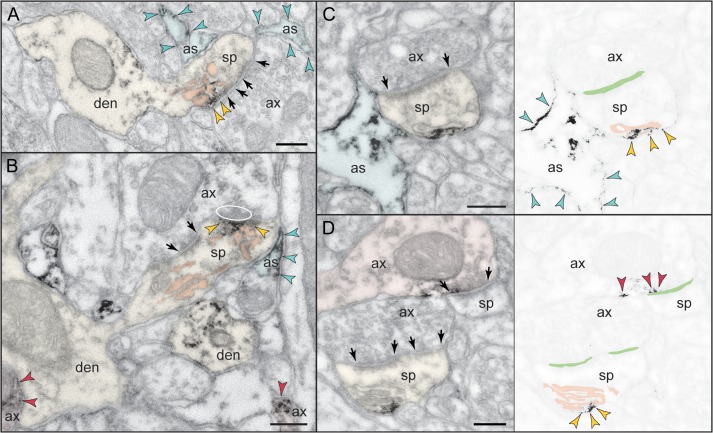
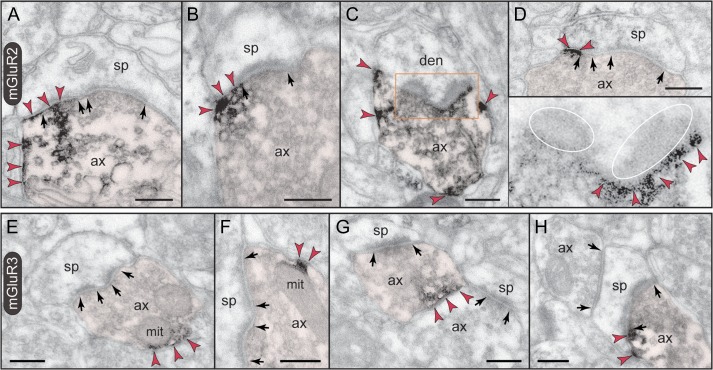
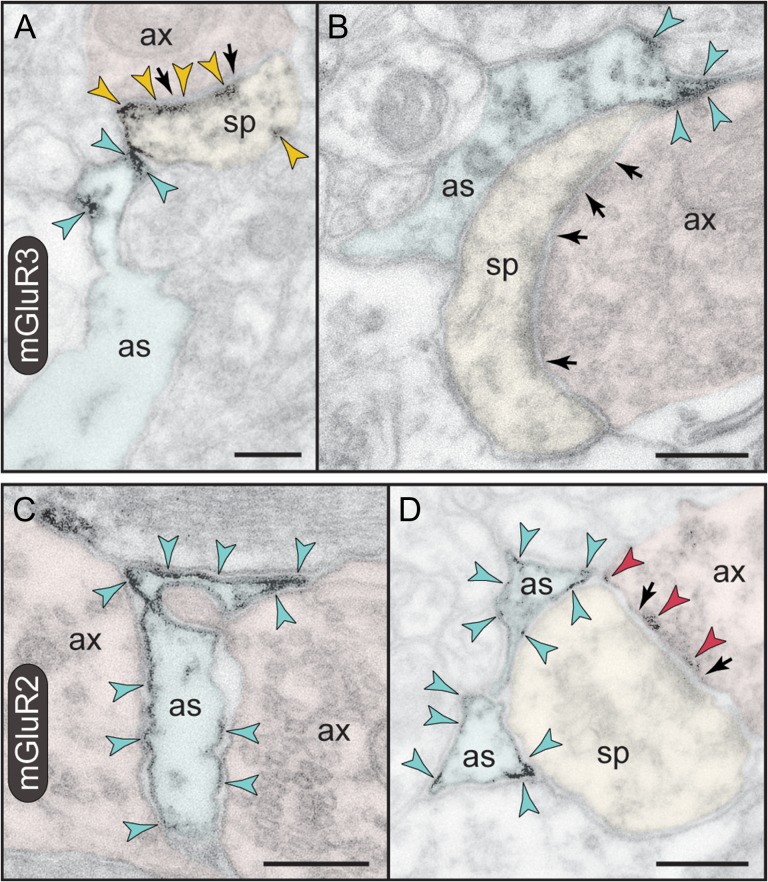
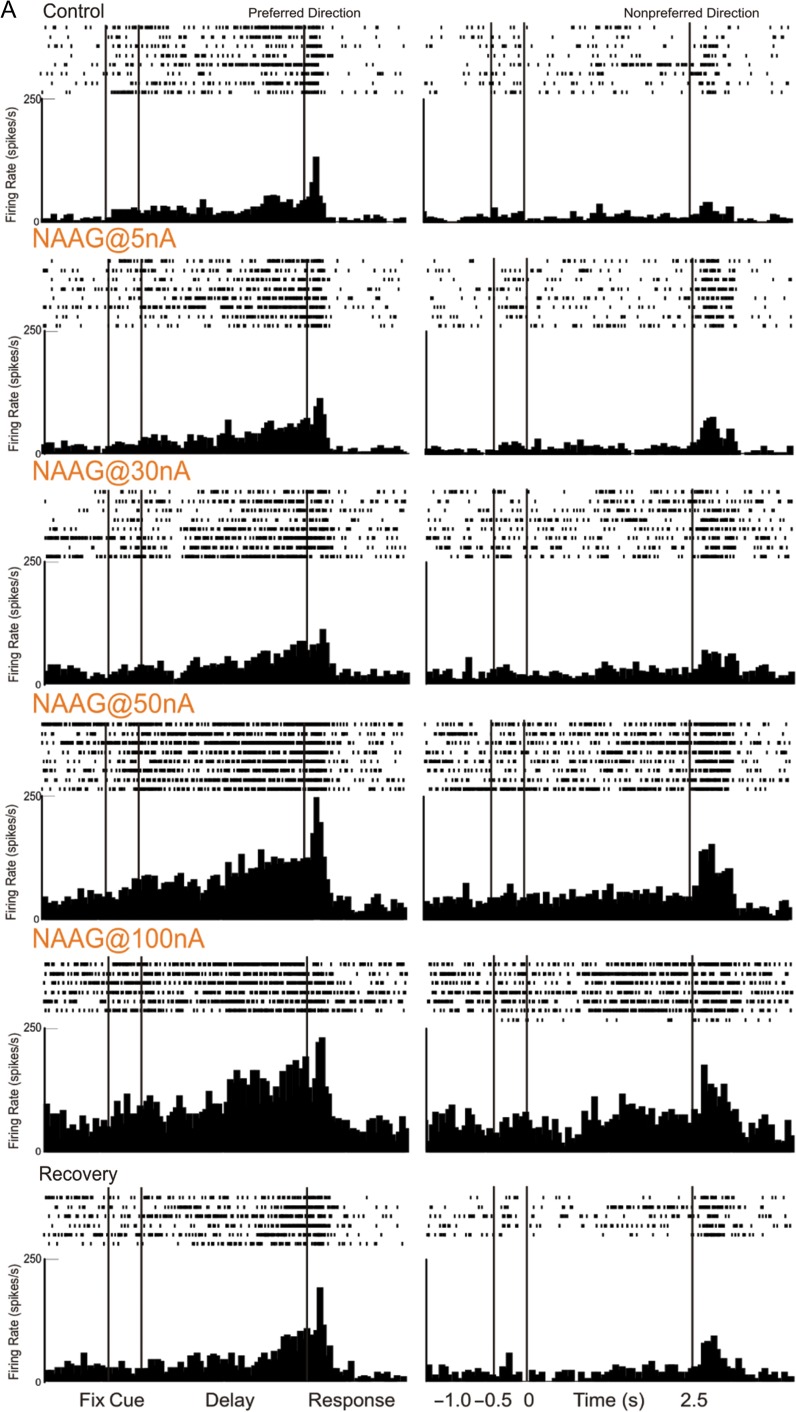
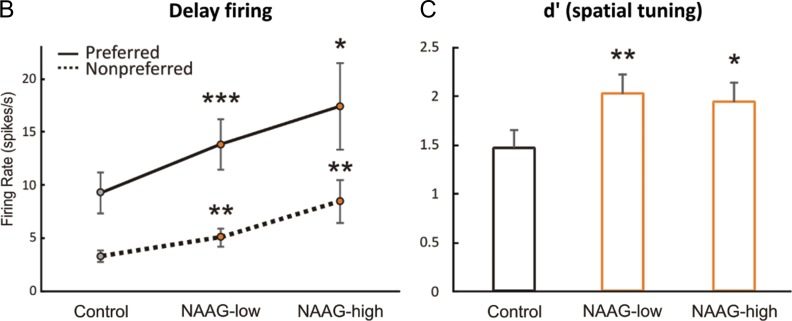
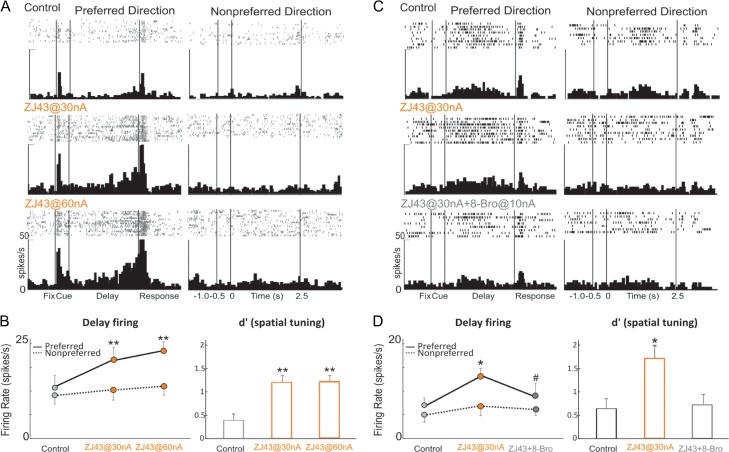
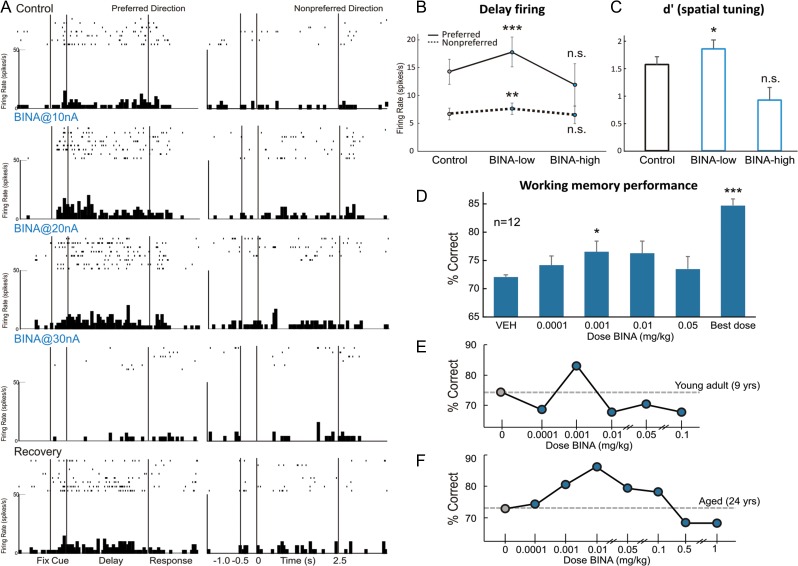
The newly evolved circuits in layer III of primate dorsolateral prefrontal cortex (dlPFC) generate the neural representations that subserve working memory. These circuits are weakened by increased cAMP-K+ channel signaling, and are a focus of pathology in schizophrenia, aging, and Alzheimer's disease. Cognitive deficits in these disorders are increasingly associated with insults to mGluR3 metabotropic glutamate receptors, while reductions in mGluR2 appear protective. This has been perplexing, as mGluR3 has been considered glial receptors, and mGluR2 and mGluR3 have been thought to have similar functions, reducing glutamate transmission. We have discovered that, in addition to their astrocytic expression, mGluR3 is concentrated postsynaptically in spine synapses of layer III dlPFC, positioned to strengthen connectivity by inhibiting postsynaptic cAMP-K+ channel actions. In contrast, mGluR2 is principally presynaptic as expected, with only a minor postsynaptic component. Functionally, increase in the endogenous mGluR3 agonist, N-acetylaspartylglutamate, markedly enhanced dlPFC Delay cell firing during a working memory task via inhibition of cAMP signaling, while the mGluR2 positive allosteric modulator, BINA, produced an inverted-U dose-response on dlPFC Delay cell firing and working memory performance. These data illuminate why insults to mGluR3 would erode cognitive abilities, and support mGluR3 as a novel therapeutic target for higher cognitive disorders.
Keywords: Alzheimer's disease; GRM2; GRM3; dendritic spine; schizophrenia.
© The Author 2017. Published by Oxford University Press. All rights reserved. For Permissions, please e-mail: journals.permissions@oup.com.
Figures
Similar articles
-
mGluR2/3 mechanisms in primate dorsolateral prefrontal cortex: evidence for both presynaptic and postsynaptic actions.Mol Psychiatry. 2017 Nov;22(11):1615-1625. doi: 10.1038/mp.2016.129. Epub 2016 Aug 9. Mol Psychiatry. 2017. PMID: 27502475 Free PMC article.
-
LY395756, an mGluR2 agonist and mGluR3 antagonist, enhances NMDA receptor expression and function in the normal adult rat prefrontal cortex, but fails to improve working memory and reverse MK801-induced working memory impairment.Exp Neurol. 2015 Nov;273:190-201. doi: 10.1016/j.expneurol.2015.08.019. Epub 2015 Sep 1. Exp Neurol. 2015. PMID: 26341392 Free PMC article.
-
Effects of blocking mGluR5 on primate dorsolateral prefrontal cortical neuronal firing and working memory performance.Psychopharmacology (Berl). 2021 Jan;238(1):97-106. doi: 10.1007/s00213-020-05661-2. Epub 2020 Sep 16. Psychopharmacology (Berl). 2021. PMID: 32939596 Free PMC article.
-
Stress and Inflammation Target Dorsolateral Prefrontal Cortex Function: Neural Mechanisms Underlying Weakened Cognitive Control.Biol Psychiatry. 2025 Feb 15;97(4):359-371. doi: 10.1016/j.biopsych.2024.06.016. Epub 2024 Jun 27. Biol Psychiatry. 2025. PMID: 38944141 Review.
-
Unusual Molecular Regulation of Dorsolateral Prefrontal Cortex Layer III Synapses Increases Vulnerability to Genetic and Environmental Insults in Schizophrenia.Biol Psychiatry. 2022 Sep 15;92(6):480-490. doi: 10.1016/j.biopsych.2022.02.003. Epub 2022 Feb 12. Biol Psychiatry. 2022. PMID: 35305820 Free PMC article. Review.
Cited by
-
A one-step protocol to generate impermeable fluorescent HaloTag substrates for in situ live cell application and super-resolution imaging.bioRxiv [Preprint]. 2024 Sep 23:2024.09.20.614087. doi: 10.1101/2024.09.20.614087. bioRxiv. 2024. PMID: 39386703 Free PMC article. Preprint.
-
Glutamate Metabotropic Receptor Type 3 (mGlu3) Localization in the Rat Prelimbic Medial Prefrontal Cortex.Front Neuroanat. 2022 Apr 4;16:849937. doi: 10.3389/fnana.2022.849937. eCollection 2022. Front Neuroanat. 2022. PMID: 35444520 Free PMC article.
-
Metabotropic glutamate receptor subtype 3 gates acute stress-induced dysregulation of amygdalo-cortical function.Mol Psychiatry. 2019 Jun;24(6):916-927. doi: 10.1038/s41380-017-0015-z. Epub 2017 Dec 21. Mol Psychiatry. 2019. PMID: 29269844 Free PMC article.
-
Progressive Transcriptional Changes in the Amygdala Implicate Neuroinflammation in the Effects of Repetitive Low-Level Blast Exposure in Male Rats.J Neurotrauma. 2023 Mar;40(5-6):561-577. doi: 10.1089/neu.2022.0282. Epub 2022 Dec 13. J Neurotrauma. 2023. PMID: 36262047 Free PMC article.
-
Effect of antipsychotic drugs on group II metabotropic glutamate receptor expression and epigenetic control in postmortem brains of schizophrenia subjects.Transl Psychiatry. 2024 Feb 23;14(1):113. doi: 10.1038/s41398-024-02832-z. Transl Psychiatry. 2024. PMID: 38396013 Free PMC article.
References
-
- Aronica E, Gorter JA, Ijlst-Keizers H, Rozemuller AJ, Yankaya B, Leenstra S, Troost D. 2003. Expression and functional role of mGluR3 and mGluR5 in human astrocytes and glioma cells: opposite regulation of glutamate transporter proteins. Eur J Neurosci. 17:2106–2118. - PubMed
-
- Benneyworth MA, Xiang Z, Smith RL, Garcia EE, Conn PJ, Sanders-Bush E. 2007. A selective positive allosteric modulator of metabotropic glutamate receptor subtype 2 blocks a hallucinogenic drug model of psychosis. Mol Pharmacol. 72:477–484. - PubMed
-
- Bishop JR, Reilly JL, Harris MS, Patel SR, Kittles R, Badner JA, Prasad KM, Nimgaonkar VL, Keshavan MS, Sweeney JA. 2015. Pharmacogenetic associations of the type-3 metabotropic glutamate receptor (GRM3) gene with working memory and clinical symptom response to antipsychotics in first-episode schizophrenia. Psychopharmacology. 232:145–154. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
