

Photosynthesis in C3-C4 intermediate Moricandia species
- PMID: 28110276
- PMCID: PMC5853546
- DOI: 10.1093/jxb/erw391
Photosynthesis in C3-C4 intermediate Moricandia species
Abstract
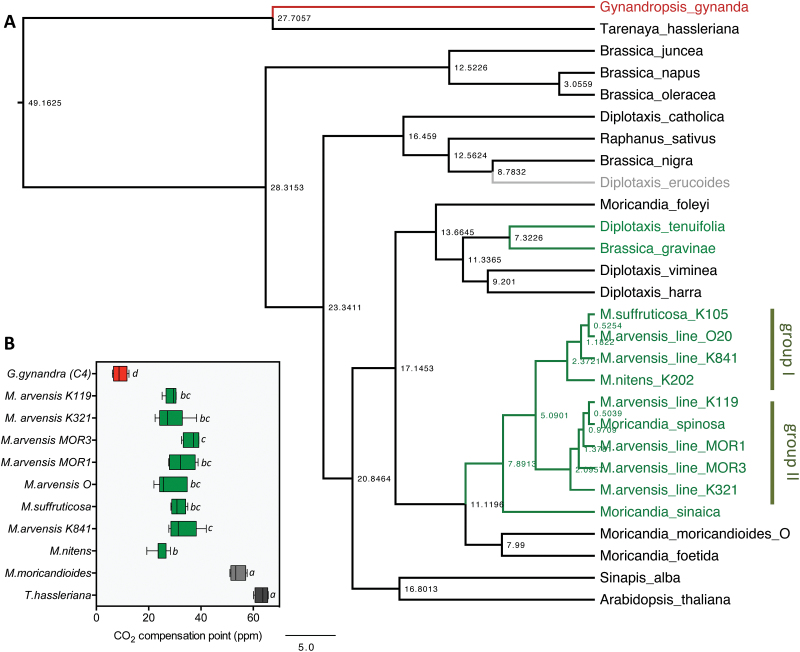
Evolution of C4 photosynthesis is not distributed evenly in the plant kingdom. Particularly interesting is the situation in the Brassicaceae, because the family contains no C4 species, but several C3-C4 intermediates, mainly in the genus Moricandia Investigation of leaf anatomy, gas exchange parameters, the metabolome, and the transcriptome of two C3-C4 intermediate Moricandia species, M. arvensis and M. suffruticosa, and their close C3 relative M. moricandioides enabled us to unravel the specific C3-C4 characteristics in these Moricandia lines. Reduced CO2 compensation points in these lines were accompanied by anatomical adjustments, such as centripetal concentration of organelles in the bundle sheath, and metabolic adjustments, such as the balancing of C and N metabolism between mesophyll and bundle sheath cells by multiple pathways. Evolution from C3 to C3-C4 intermediacy was probably facilitated first by loss of one copy of the glycine decarboxylase P-protein, followed by dominant activity of a bundle sheath-specific element in its promoter. In contrast to recent models, installation of the C3-C4 pathway was not accompanied by enhanced activity of the C4 cycle. Our results indicate that metabolic limitations connected to N metabolism or anatomical limitations connected to vein density could have constrained evolution of C4 in Moricandia.
Keywords: Moricandia.; Bundle sheath; C3–C4 intermediacy; C4 photosynthesis; evolution; glycine decarboxylase.
© The Author 2016. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures







References
-
- Adwy W, Laxa M, Peterhansel C. 2015. A simple mechanism for the establishment of C2-specific gene expression in Brassicaceae. The Plant Journal 84, 1231–1238. - PubMed
-
- Apel P, Horstmann C, Pfeffer M. 1997. The Moricandia syndrome in species of the Brassicaceae – evolutionary aspects. Photosynthetica 33, 205–215.
-
- Arias T, Beilstein MA, Tang M, McKain MR, Pires JC. 2014. Diversification times, among Brassica (Brassicaceae) crops suggest hybrid formation after 20 million years of divergence. American Journal of Botany 101, 86–91. - PubMed
-
- Ashton AR, Burnell JN, Furbank RT, Jenkins CLD, Hatch MD. 1990. The enzymes in C4 photosynthesis. In: Lea PJ, Harborne JB, eds. Enzymes of primary metabolism. London, UK: Academic Press, 39–72.
-
- Aubry S, Brown NJ, Hibberd JM. 2011. The role of proteins in C3 plants prior to their recruitment into the C4 pathway. Journal of Experimantal Botany 62, 3049–3059. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
