

A GluD Coming-Of-Age Story
- PMID: 28110935
- PMCID: PMC5553105
- DOI: 10.1016/j.tins.2016.12.004
A GluD Coming-Of-Age Story
Abstract
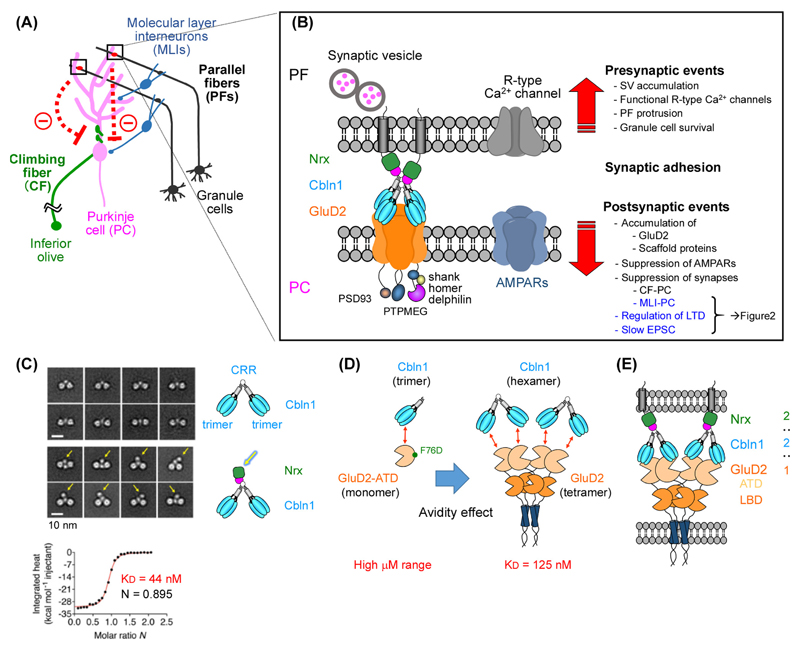
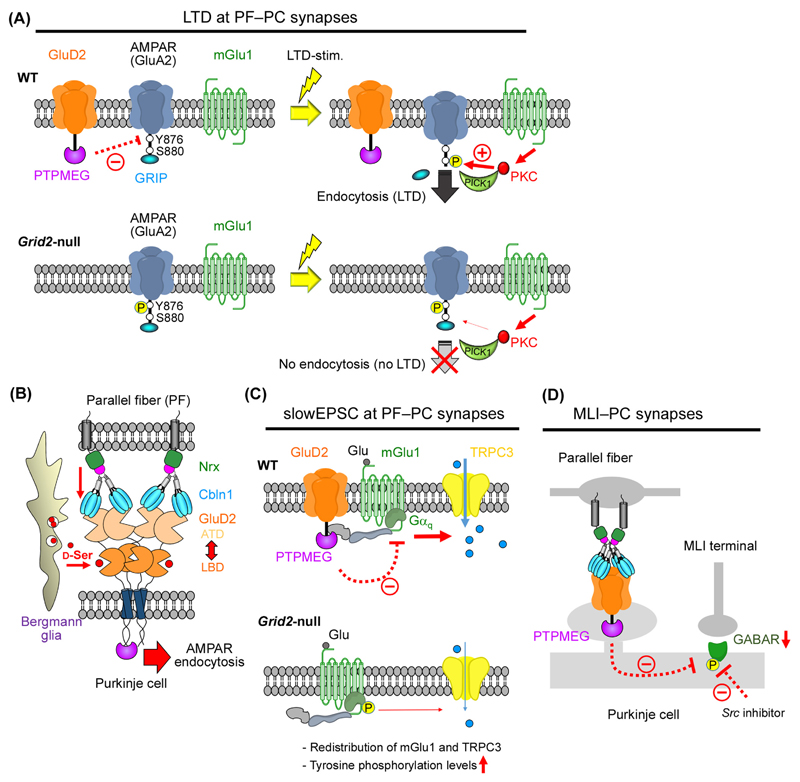
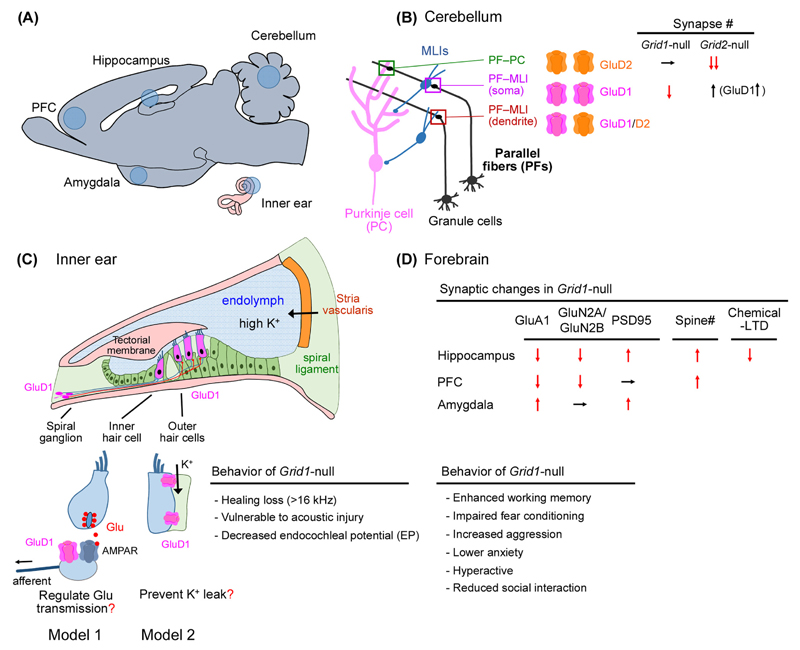
The GluD1 and GluD2 receptors form the GluD ionotropic glutamate receptor (iGluR) subfamily. Without known endogenous ligands, they have long been referred to as 'orphan' and remained enigmatic functionally. Recent progress has, however, radically changed this view. Both GluD receptors are expressed in wider brain regions than originally thought. Human genetic studies and analyses of knockout mice have revealed their involvement in multiple neurodevelopmental and psychiatric disorders. The discovery of endogenous ligands, together with structural investigations, has opened the way towards a mechanistic understanding of GluD signaling at central nervous system synapses. These studies have also prompted the hypothesis that all iGluRs, and potentially other neurotransmitter receptors, rely on the cooperative binding of extracellular small-molecule and protein ligands for physiological signaling.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures
References
-
- Yuzaki M. The delta2 glutamate receptor: 10 years later. Neurosci Res. 2003;46:11–22. - PubMed
-
- Hirano T. Cerebellar regulation mechanisms learned from studies on GluRδ2. Mol Neurobiol. 2006;33:1–15. - PubMed
-
- Mandolesi G, et al. An orphan ionotropic glutamate receptor: the delta2 subunit. Neuroscience. 2009;158:67–77. - PubMed
-
- Schmid SM, Hollmann M. Bridging the synaptic cleft: lessons from orphan glutamate receptors. Sci Signal. 2010;3:pe28. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
