

Interdomain Communication of the Chd1 Chromatin Remodeler across the DNA Gyres of the Nucleosome
- PMID: 28111016
- PMCID: PMC5308885
- DOI: 10.1016/j.molcel.2016.12.011
Interdomain Communication of the Chd1 Chromatin Remodeler across the DNA Gyres of the Nucleosome
Abstract
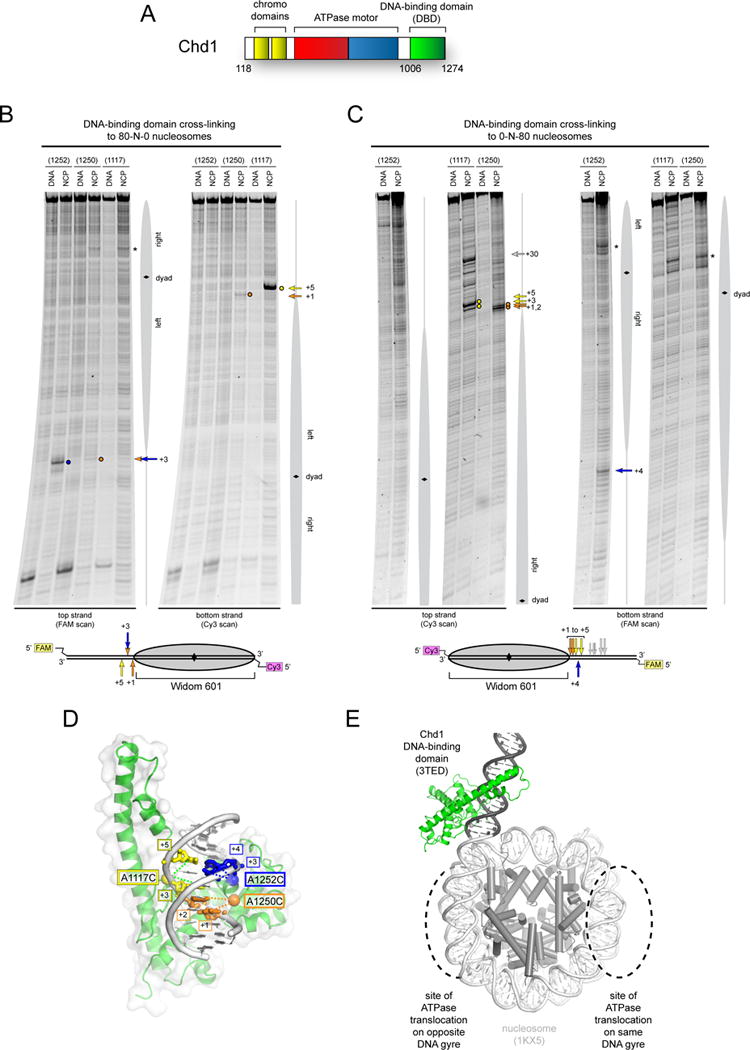
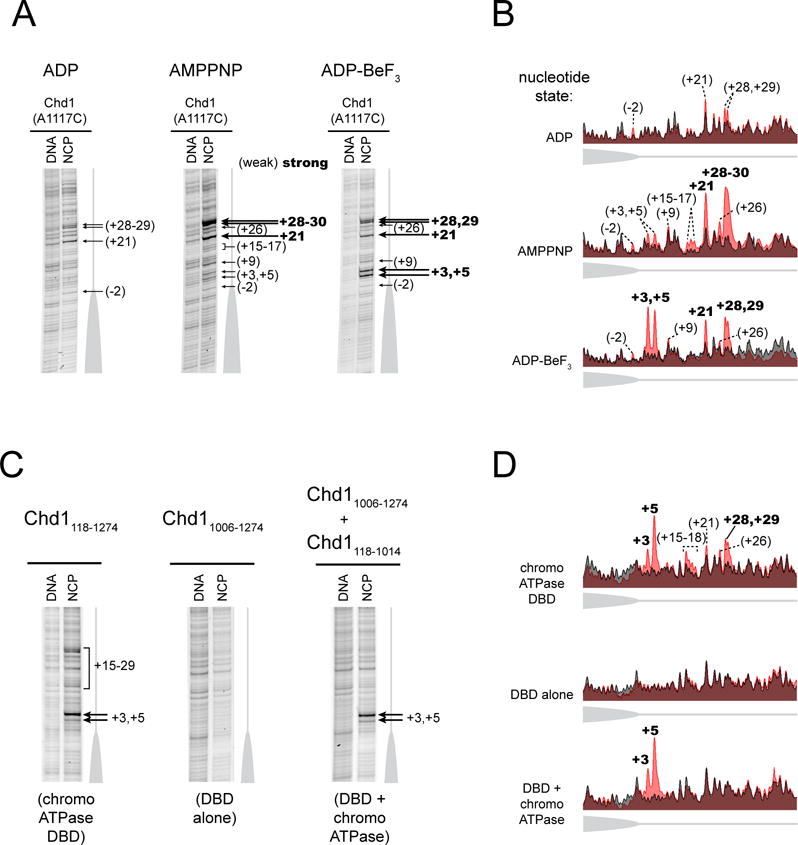
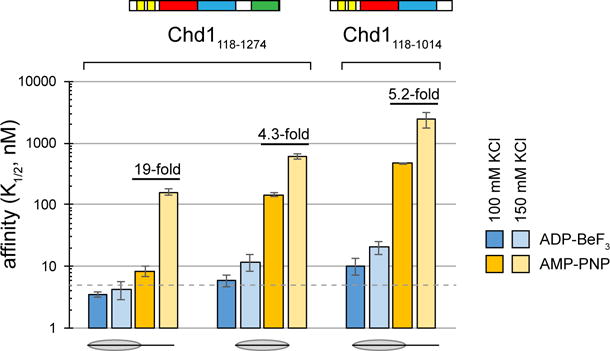
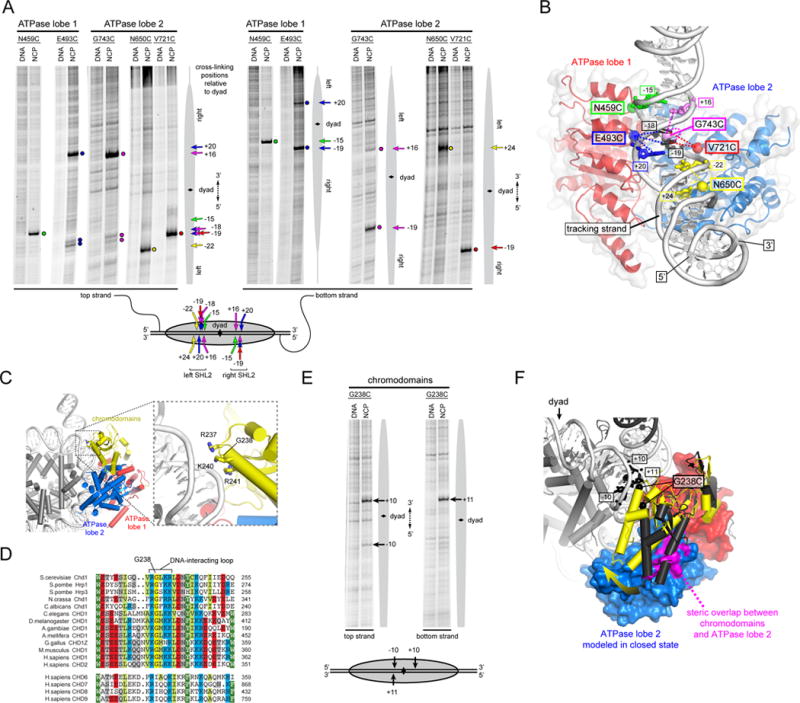
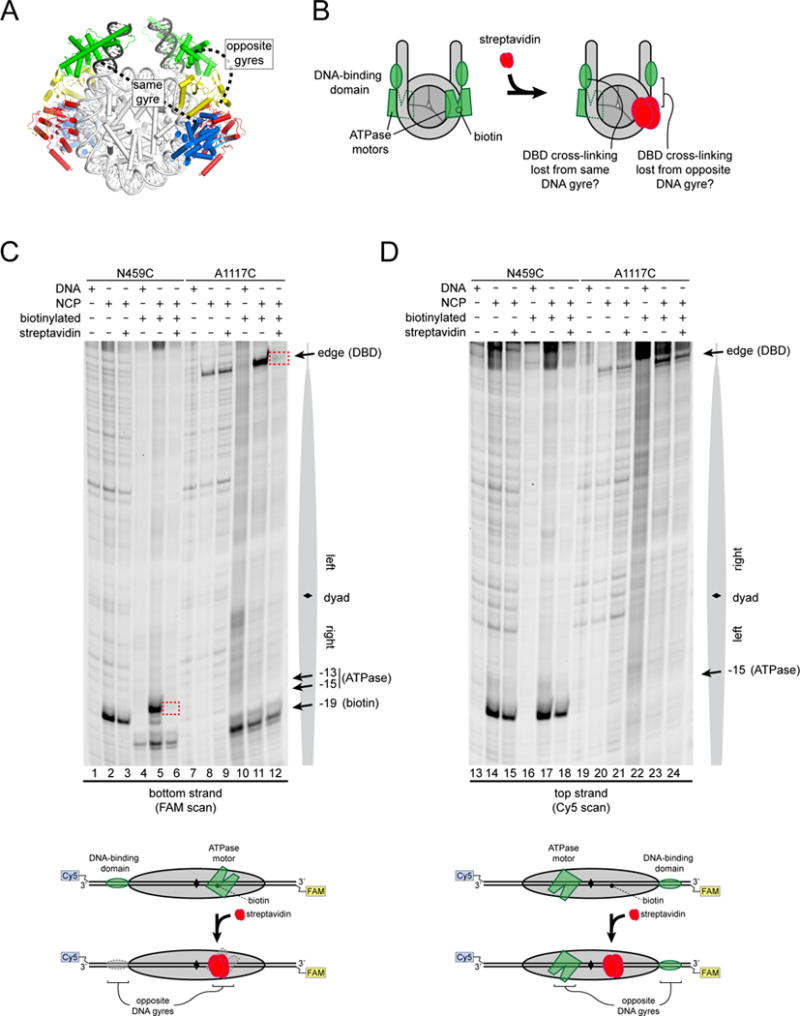
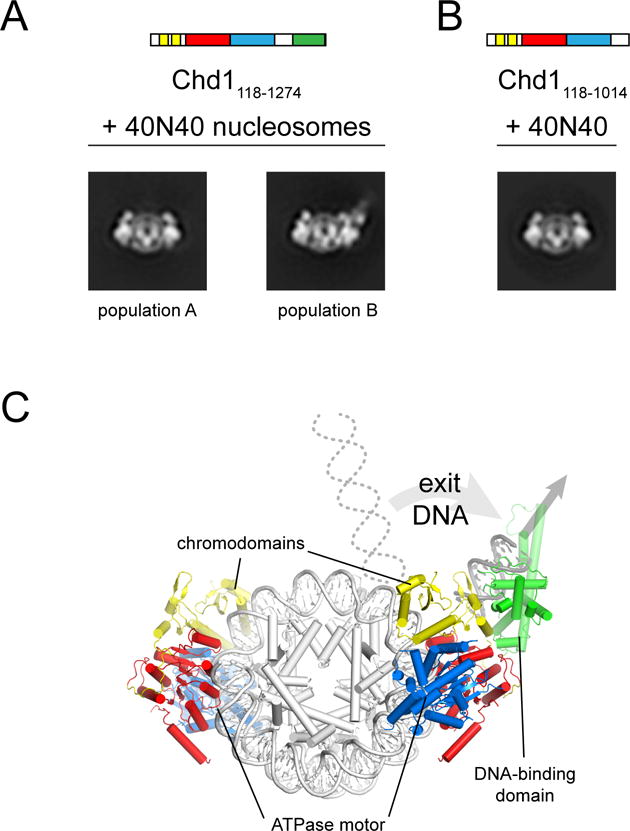
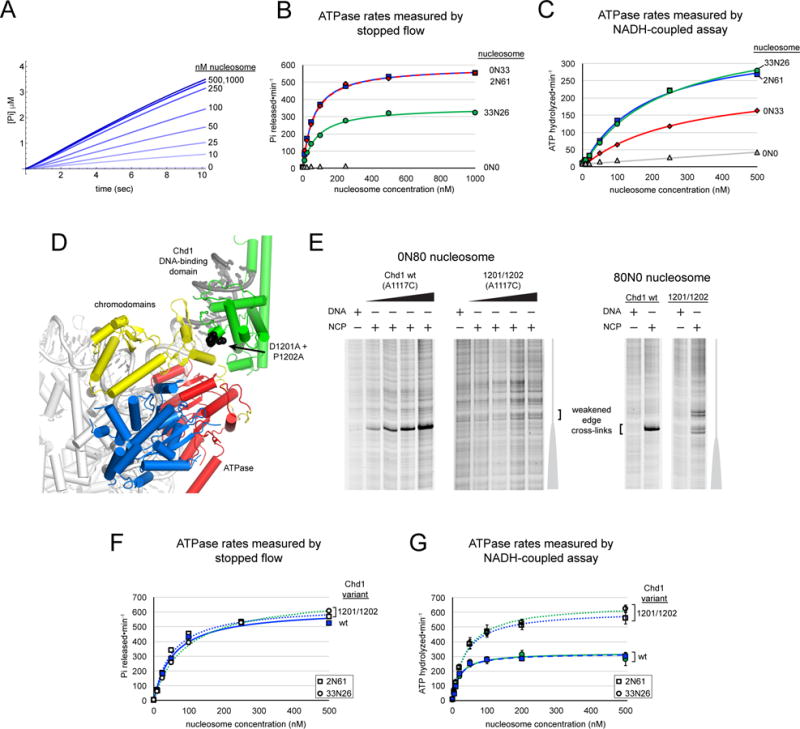
Chromatin remodelers use a helicase-like ATPase motor to reposition and reorganize nucleosomes along genomic DNA. Yet, how the ATPase motor communicates with other remodeler domains in the context of the nucleosome has so far been elusive. Here, we report for the Chd1 remodeler a unique organization of domains on the nucleosome that reveals direct domain-domain communication. Site-specific cross-linking shows that the chromodomains and ATPase motor bind to adjacent SHL1 and SHL2 sites, respectively, on nucleosomal DNA and pack against the DNA-binding domain on DNA exiting the nucleosome. This domain arrangement spans the two DNA gyres of the nucleosome and bridges both ends of a wrapped, ∼90-bp nucleosomal loop of DNA, suggesting a means for nucleosome assembly. This architecture illustrates how Chd1 senses DNA outside the nucleosome core and provides a basis for nucleosome spacing and directional sliding away from transcription factor barriers.
Keywords: CHD remodeler; DNA unwrapping; ISWI remodeler; SF2 ATPase; SHL2; chromatin remodeling; chromodomains; nucleosome assembly; nucleosome sliding; superhelical location 2.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Brune M, Hunter JL, Corrie JE, Webb MR. Direct, real-time measurement of rapid inorganic phosphate release using a novel fluorescent probe and its application to actomyosin subfragment 1 ATPase. Biochemistry. 1994;27:8262–8271. - PubMed
-
- Davey CA, Sargent DF, Luger K, Maeder AW, Richmond TJ. Solvent mediated interactions in the structure of the nucleosome core particle at 1.9 a resolution. J Mol Biol. 2002;5:1097–1113. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
