

Role of Mitochondrial Metabolism in the Control of Early Lineage Progression and Aging Phenotypes in Adult Hippocampal Neurogenesis
- PMID: 28111078
- PMCID: PMC5300896
- DOI: 10.1016/j.neuron.2016.12.017
Role of Mitochondrial Metabolism in the Control of Early Lineage Progression and Aging Phenotypes in Adult Hippocampal Neurogenesis
Erratum in
-
Role of Mitochondrial Metabolism in the Control of Early Lineage Progression and Aging Phenotypes in Adult Hippocampal Neurogenesis.Neuron. 2017 Mar 22;93(6):1518. doi: 10.1016/j.neuron.2017.03.008. Neuron. 2017. PMID: 28334613 Free PMC article. No abstract available.
Abstract
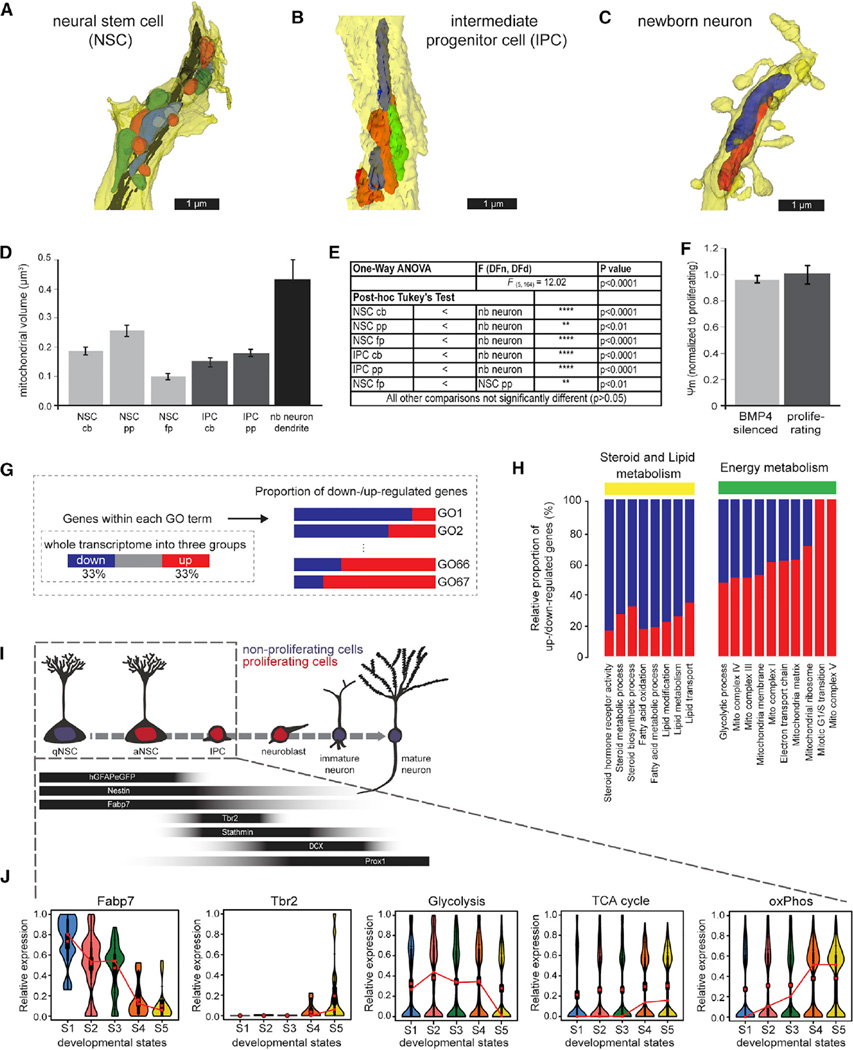
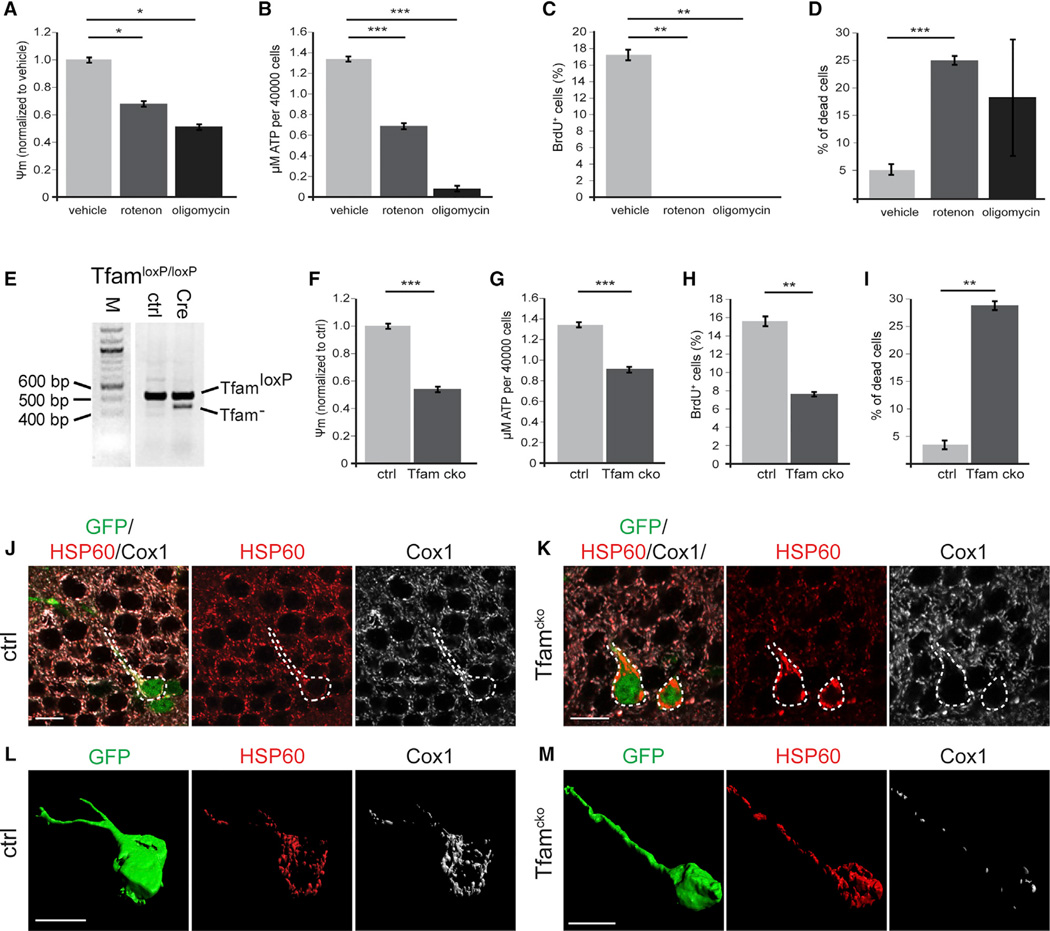
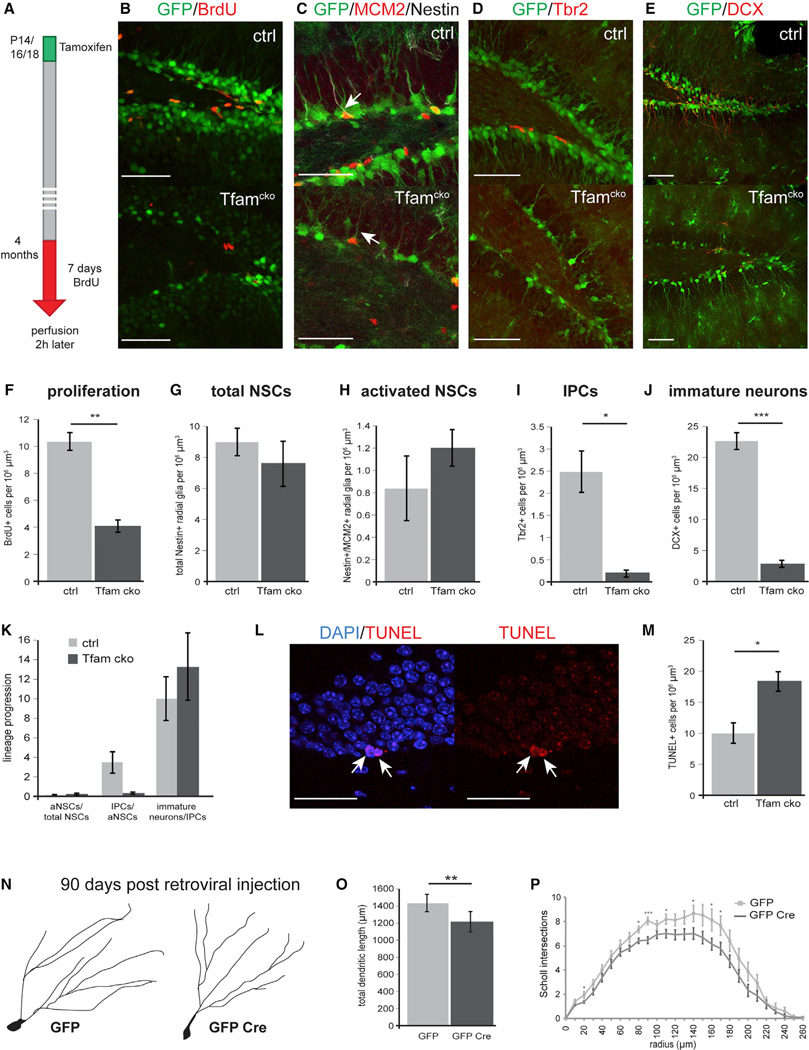
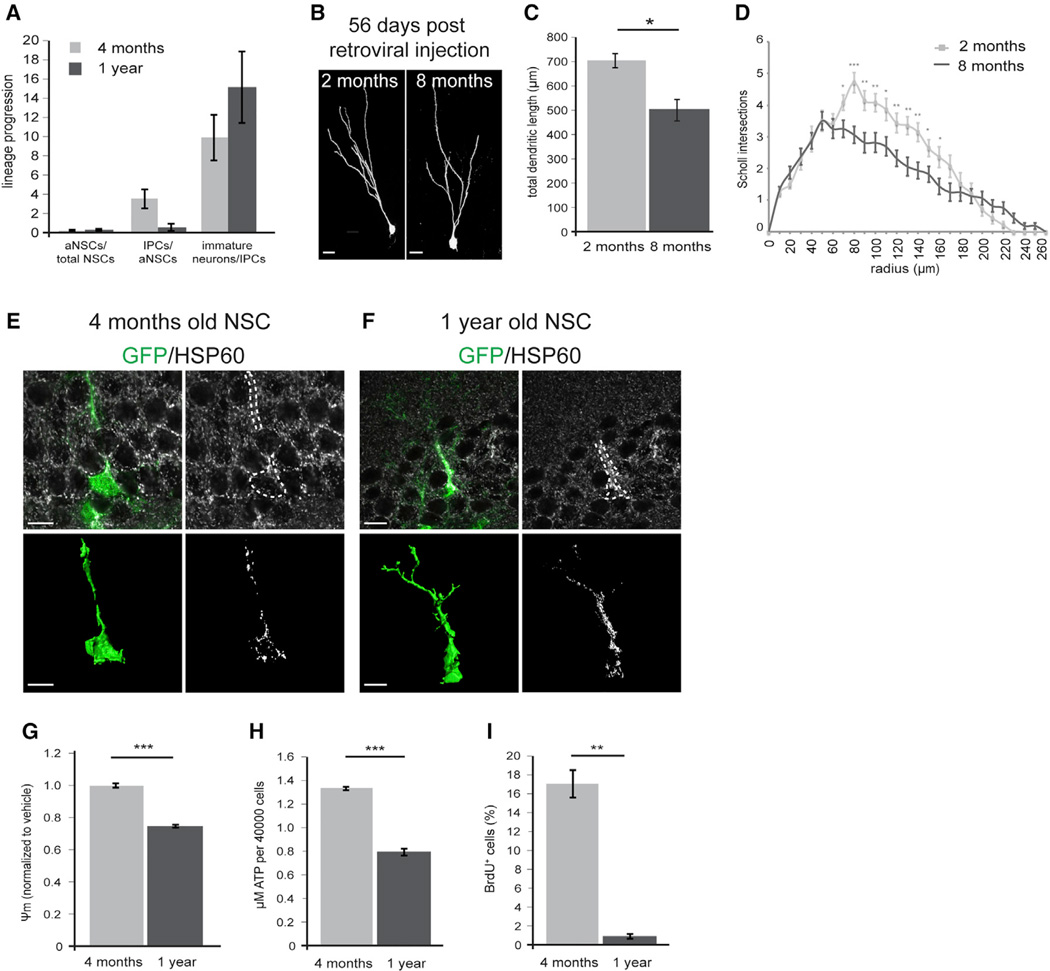
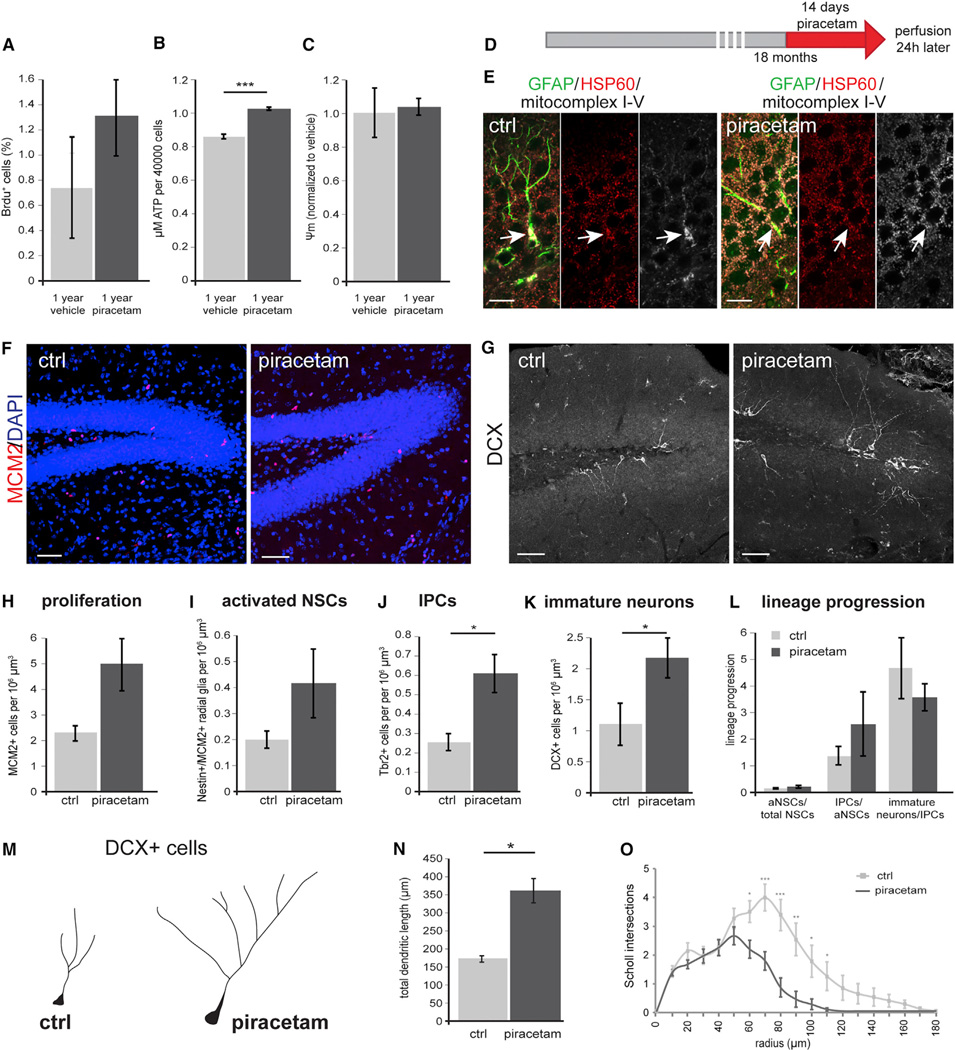
Precise regulation of cellular metabolism is hypothesized to constitute a vital component of the developmental sequence underlying the life-long generation of hippocampal neurons from quiescent neural stem cells (NSCs). The identity of stage-specific metabolic programs and their impact on adult neurogenesis are largely unknown. We show that the adult hippocampal neurogenic lineage is critically dependent on the mitochondrial electron transport chain and oxidative phosphorylation machinery at the stage of the fast proliferating intermediate progenitor cell. Perturbation of mitochondrial complex function by ablation of the mitochondrial transcription factor A (Tfam) reproduces multiple hallmarks of aging in hippocampal neurogenesis, whereas pharmacological enhancement of mitochondrial function ameliorates age-associated neurogenesis defects. Together with the finding of age-associated alterations in mitochondrial function and morphology in NSCs, these data link mitochondrial complex function to efficient lineage progression of adult NSCs and identify mitochondrial function as a potential target to ameliorate neurogenesis-defects in the aging hippocampus.
Keywords: adult neurogenesis; aging; metabolism; mitochondria; stem cells.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Revealing the Hidden Powers that Fuel Adult Neurogenesis.Cell Stem Cell. 2017 Feb 2;20(2):154-156. doi: 10.1016/j.stem.2017.01.004. Cell Stem Cell. 2017. PMID: 28157496
References
-
- Ahlqvist KJ, Hämälaänen RH, Yatsuga S, Uutela M, Terzioglu M, Götz A, Forsström S, Salven P, Angers-Loustau A, Kopra OH, et al. Somatic progenitor cell vulnerability to mitochondrial DNA mutagenesis underlies progeroid phenotypes in Polg mutator mice. Cell Metab. 2012;15:100–109. - PubMed
-
- Alirol E, Martinou JC. Mitochondria and cancer: is there a morphological connection? Oncogene. 2006;25:4706–4716. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
