

SUMO-modified insulin-like growth factor 1 receptor (IGF-1R) increases cell cycle progression and cell proliferation
- PMID: 28112398
- PMCID: PMC5518303
- DOI: 10.1002/jcp.25818
SUMO-modified insulin-like growth factor 1 receptor (IGF-1R) increases cell cycle progression and cell proliferation
Abstract
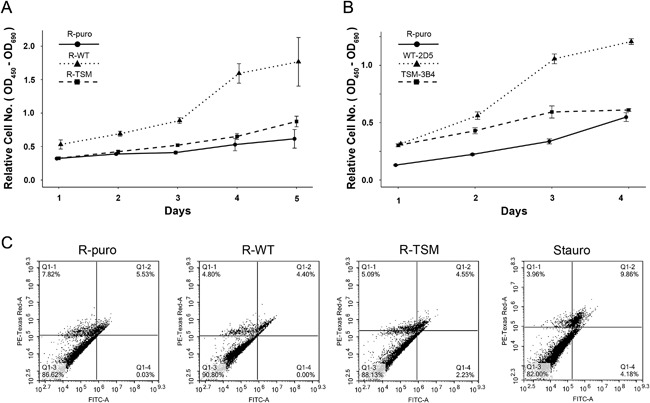
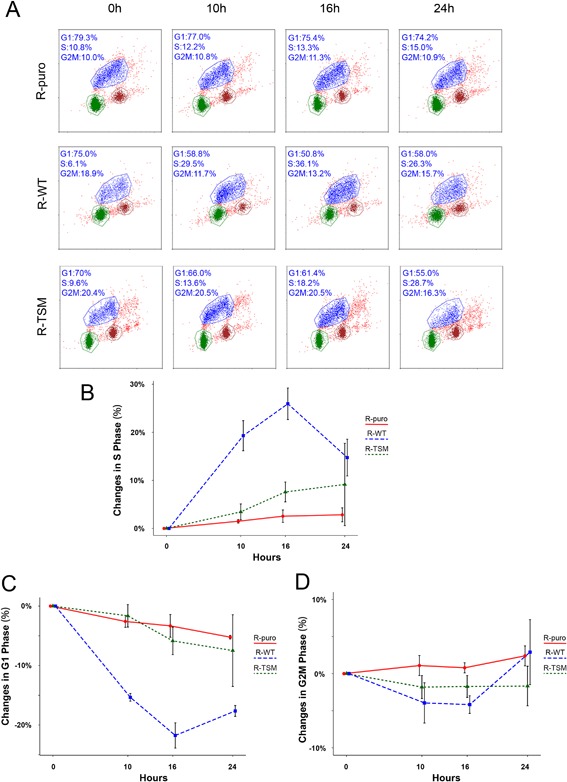
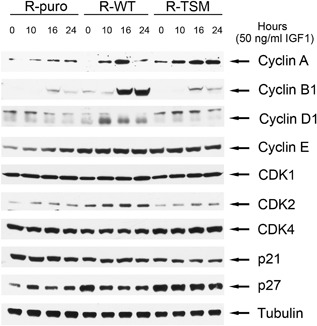
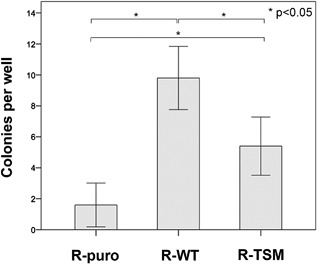
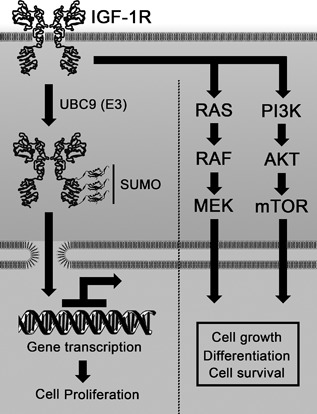
Increasing number of studies have shown nuclear localization of the insulin-like growth factor 1 receptor (nIGF-1R) in tumor cells and its links to adverse clinical outcome in various cancers. Any obvious cell physiological roles of nIGF-1R have, however, still not been disclosed. Previously, we reported that IGF-1R translocates to cell nucleus and modulates gene expression by binding to enhancers, provided that the receptor is SUMOylated. In this study, we constructed stable transfectants of wild type IGF1R (WT) and triple-SUMO-site-mutated IGF1R (TSM) using igf1r knockout mouse fibroblasts (R-). Cell clones (R-WT and R-TSM) expressing equal amounts of IGF-1R were selected for experiments. Phosphorylation of IGF-1R, Akt, and Erk upon IGF-1 stimulation was equal in R-WT and R-TSM. WT was confirmed to enter nuclei. TSM did also undergo nuclear translocation, although to a lesser extent. This may be explained by that TSM heterodimerizes with insulin receptor, which is known to translocate to cell nuclei. R-WT proliferated substantially faster than R-TSM, which did not differ significantly from the empty vector control. Upon IGF-1 stimulation G1-S-phase progression of R-WT increased from 12 to 38%, compared to 13 to 20% of R-TSM. The G1-S progression of R-WT correlated with increased expression of cyclin D1, A, and CDK2, as well as downregulation of p27. This suggests that SUMO-IGF-1R affects upstream mechanisms that control and coordinate expression of cell cycle regulators. Further studies to identify such SUMO-IGF-1R dependent mechanisms seem important.
Keywords: IGF-1R; SUMOylation; cancer; cell cycle; proliferation.
© 2017 The Authors. Journal of Cellular Physiology Published by Wiley Periodicals Inc.
Figures

References
-
- Asmane, I. , Watkin, E. , Alberti, L. , Duc, A. , Marec‐Berard, P. , Ray‐Coquard, I. , … Bergerat, J. P. (2012). Insulin‐like growth factor type 1 receptor (IGF‐1R) exclusive nuclear staining: A predictive biomarker for IGF‐1R monoclonal antibody (Ab) therapy in sarcomas. European Journal of Cancer, 48, 3027–3035. - PubMed
-
- Baserga, R. (2009). The insulin receptor substrate‐1: A biomarker for cancer? Experimental Cell Research, 315, 727–732. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
