

Persistence of Coxsackievirus B4 in pancreatic ductal-like cells results in cellular and viral changes
- PMID: 28112573
- PMCID: PMC5711397
- DOI: 10.1080/21505594.2017.1284735
Persistence of Coxsackievirus B4 in pancreatic ductal-like cells results in cellular and viral changes
Abstract
Introduction: Although known as cytolytic viruses, group B coxackieviruses (CVB) are able to establish a persistent infection in vitro and in vivo. Viral persistence has been reported as a key mechanism in the pathogenesis of CVB-associated chronic diseases such as type 1 diabetes (T1D). The impact of CVB4 persistence on human pancreas ductal-like cells was investigated.
Methods: A persistent CVB4 infection was established in ductal-like cells. PDX-1 expression, resistance to CVB4-induced lysis and CAR expression were evaluated. The profile of cellular microRNAs (miRNAs) was investigated through miRNA-sequencing. Viral phenotypic changes were examined, and genomic modifications were assessed by sequencing of the viral genome.
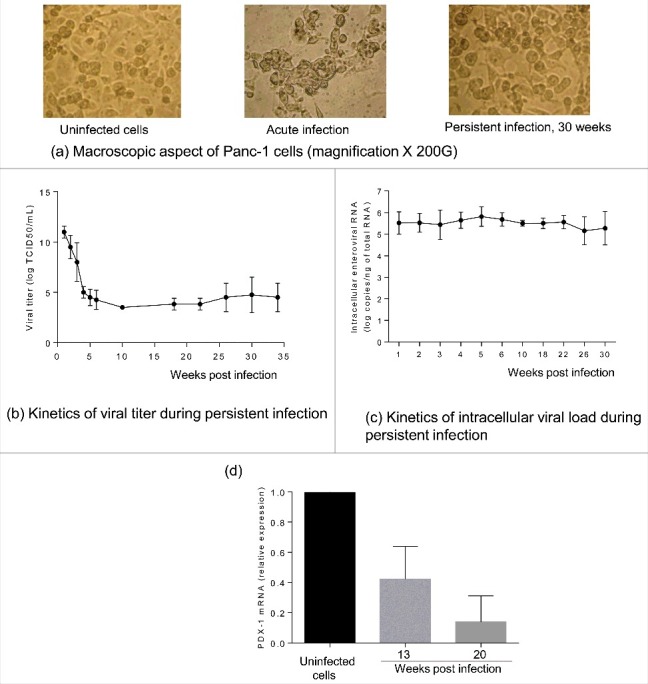
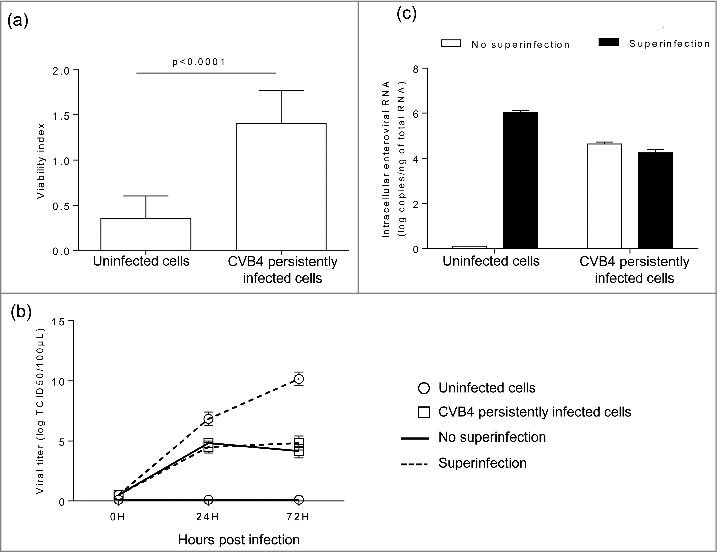
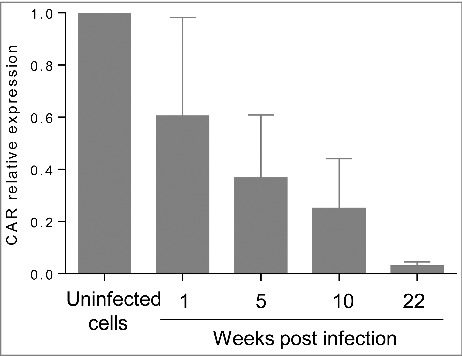
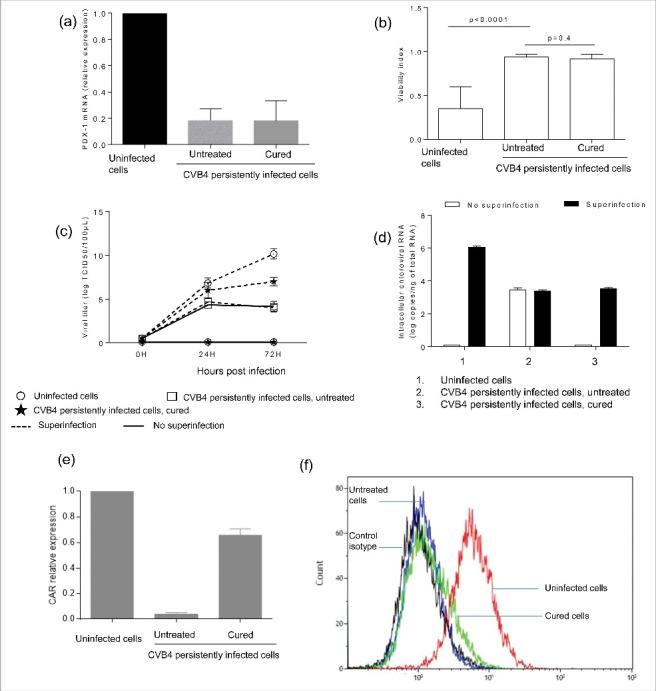
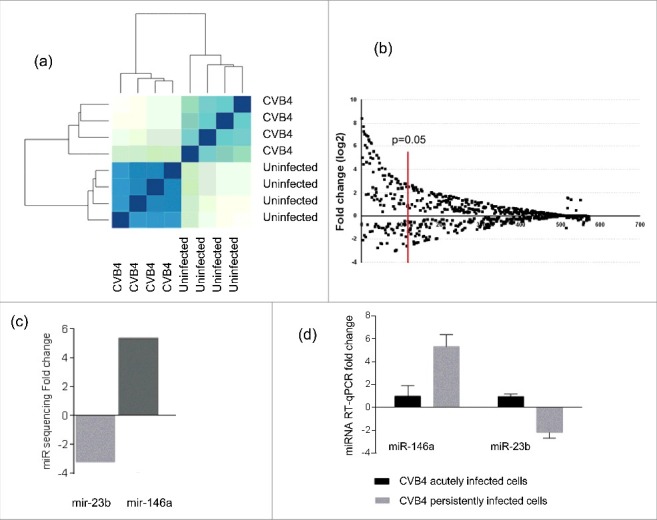
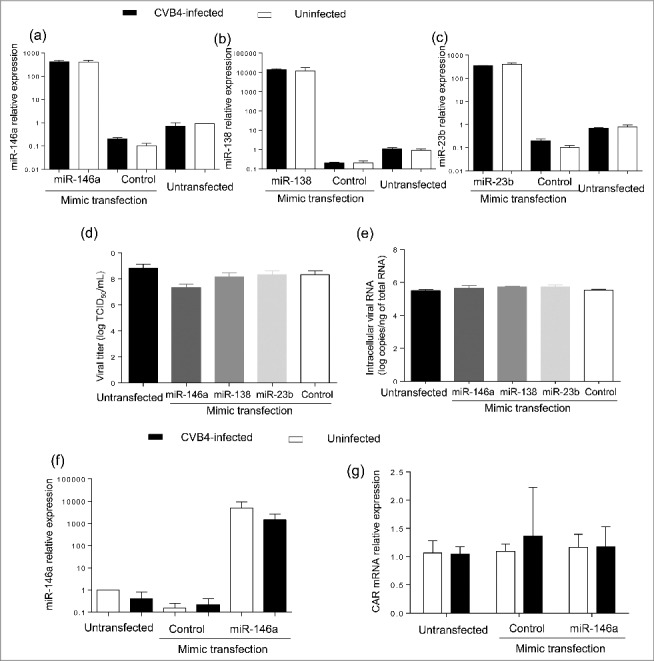
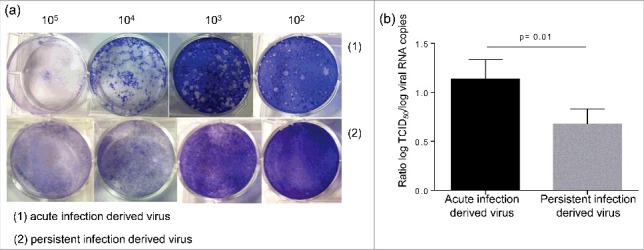
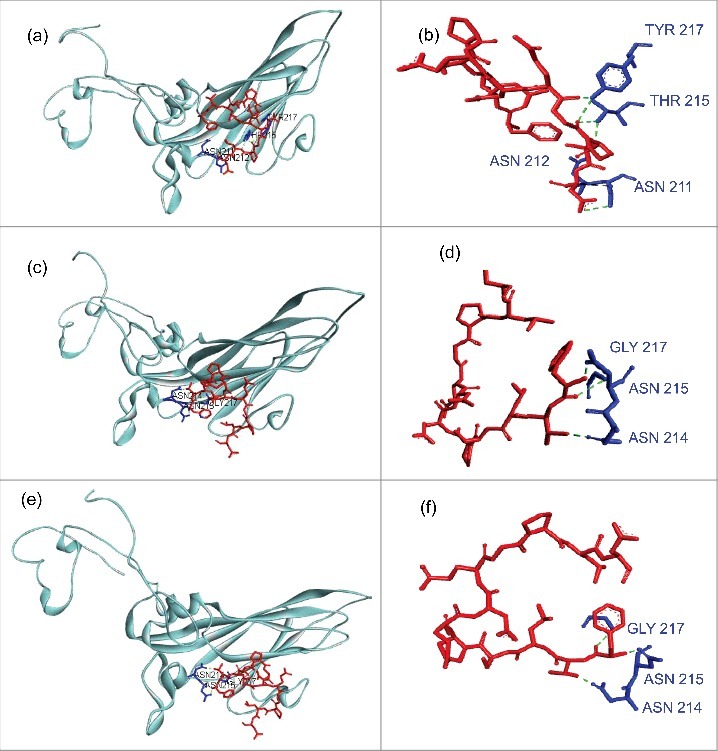
Results: The CVB4 persistence in ductal-like cells was productive, with continuous release of infectious particles. Persistently infected cells displayed a resistance to CVB4-induced lysis upon superinfection and expression of PDX-1 and CAR was decreased. These changes were maintained even after virus clearance. The patterns of cellular miRNA expression in mock-infected and in CVB4-persistently infected ductal-like cells were clearly different. The persistent infection-derived virus (PIDV) was still able to induce cytopathic effect but its plaques were smaller than the parental virus. Several mutations appeared in various PIDV genome regions, but amino acid substitutions did not affect the predicted site of interaction with CAR.
Conclusion: Cellular and viral changes occur during persistent infection of human pancreas ductal-like cells with CVB4. The persistence of cellular changes even after virus clearance supports the hypothesis of a long-lasting impact of persistent CVB infection on the cells.
Keywords: CAR; Coxsackievirus B4; PDX-1; miRNA; pancreatic cells; persistence.
Figures

Comment in
-
Acute and chronic disease caused by enteroviruses.Virulence. 2017 Oct 3;8(7):1062-1065. doi: 10.1080/21505594.2017.1308620. Epub 2017 Mar 31. Virulence. 2017. PMID: 28362547 Free PMC article. No abstract available.
References
-
- Knowles N, Hovi T, Hyypiä T. Picornaviridae. In: Virus Taxonomy: Classification and Nomenclature of Viruses: Ninth Report of the International Committee on Taxonomy of Viruses San Diego: King, A.M.Q., Adams, M.J., Carstens, E.B. and Lefkowitz, E.J; 2012. page 855-80.
-
- Tapparel C, Siegrist F, Petty TJ, Kaiser L. Picornavirus and enterovirus diversity with associated human diseases. Infect Genet Evol J Mol Epidemiol Evol Genet Infect Dis 2013; 14:282-93; http://dx.doi.org/ 10.1016/j.meegid.2012.10.016 - DOI - PubMed
-
- Romero JR. Pediatric group B coxsackievirus infections. Curr Top Microbiol Immunol 2008; 323:223-39; PMID:18357772 - PubMed
-
- Hober D, Alidjinou EK. Enteroviral pathogenesis of type 1 diabetes: queries and answers. Curr Opin Infect Dis 2013; 26:263-9; PMID:23549392; http://dx.doi.org/ 10.1097/QCO.0b013e3283608300 - DOI - PubMed
-
- Hober D, Sauter P. Pathogenesis of type 1 diabetes mellitus: interplay between enterovirus and host. Nat Rev Endocrinol 2010; 6:279-89; PMID:20351698; http://dx.doi.org/ 10.1038/nrendo.2010.27 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources