

Maturation of the infant microbiome community structure and function across multiple body sites and in relation to mode of delivery
- PMID: 28112736
- PMCID: PMC5345907
- DOI: 10.1038/nm.4272
Maturation of the infant microbiome community structure and function across multiple body sites and in relation to mode of delivery
Abstract
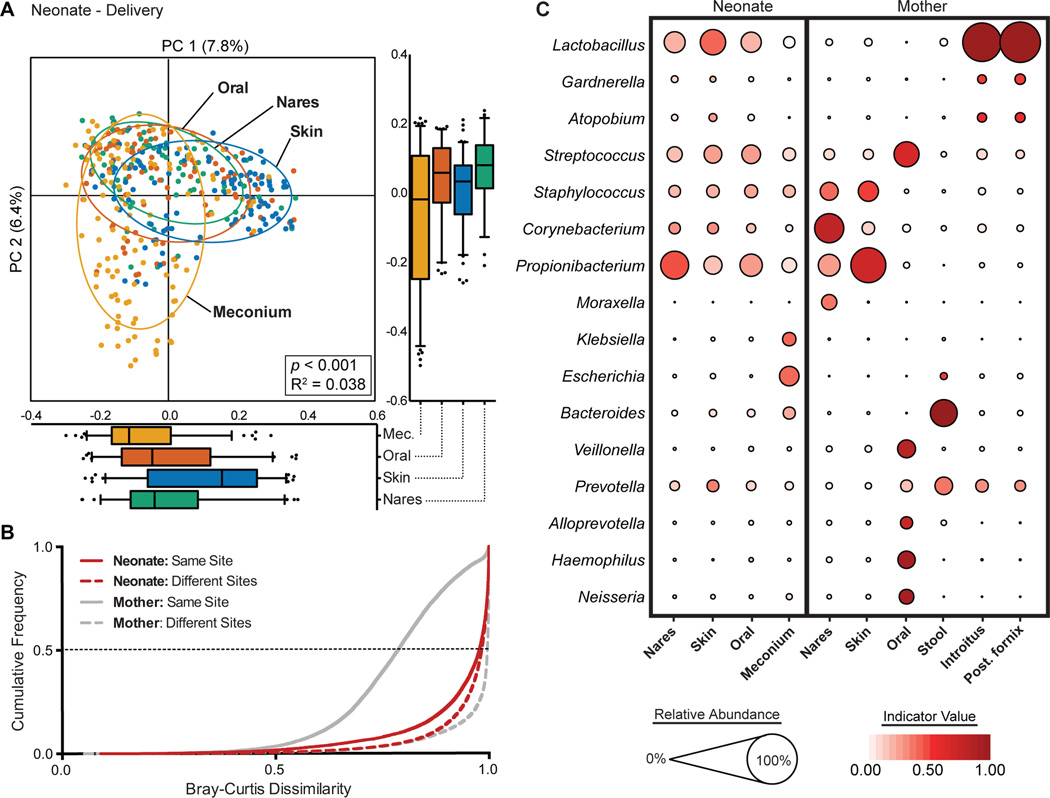
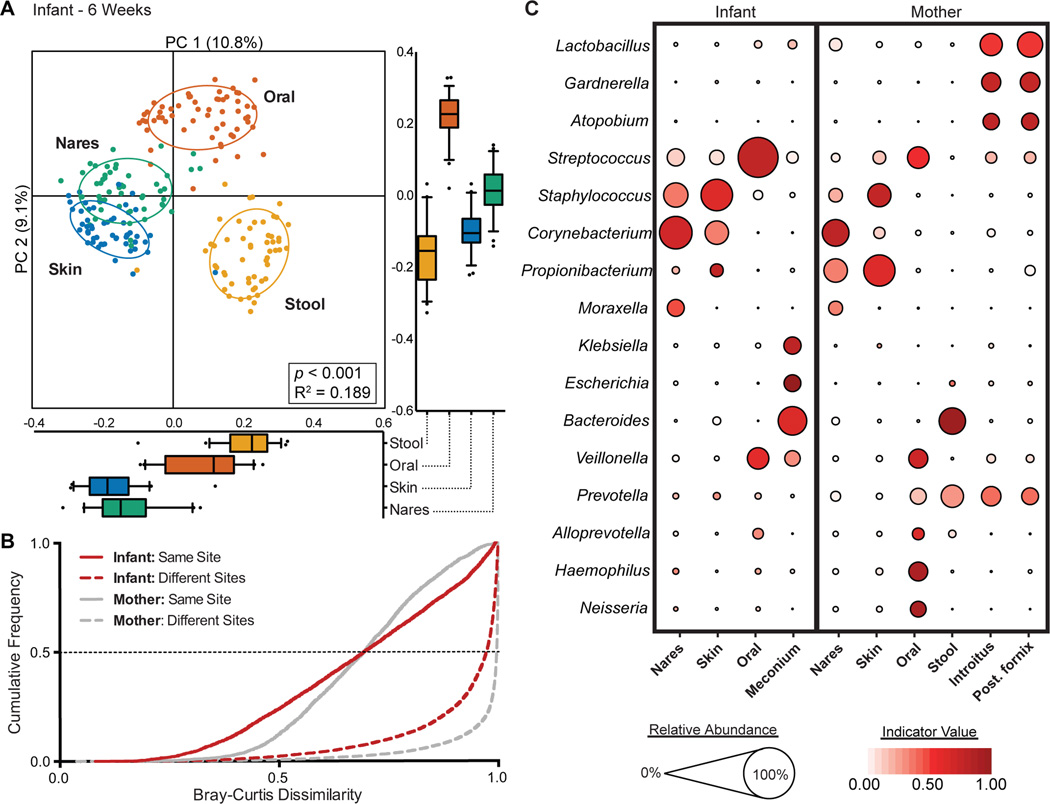
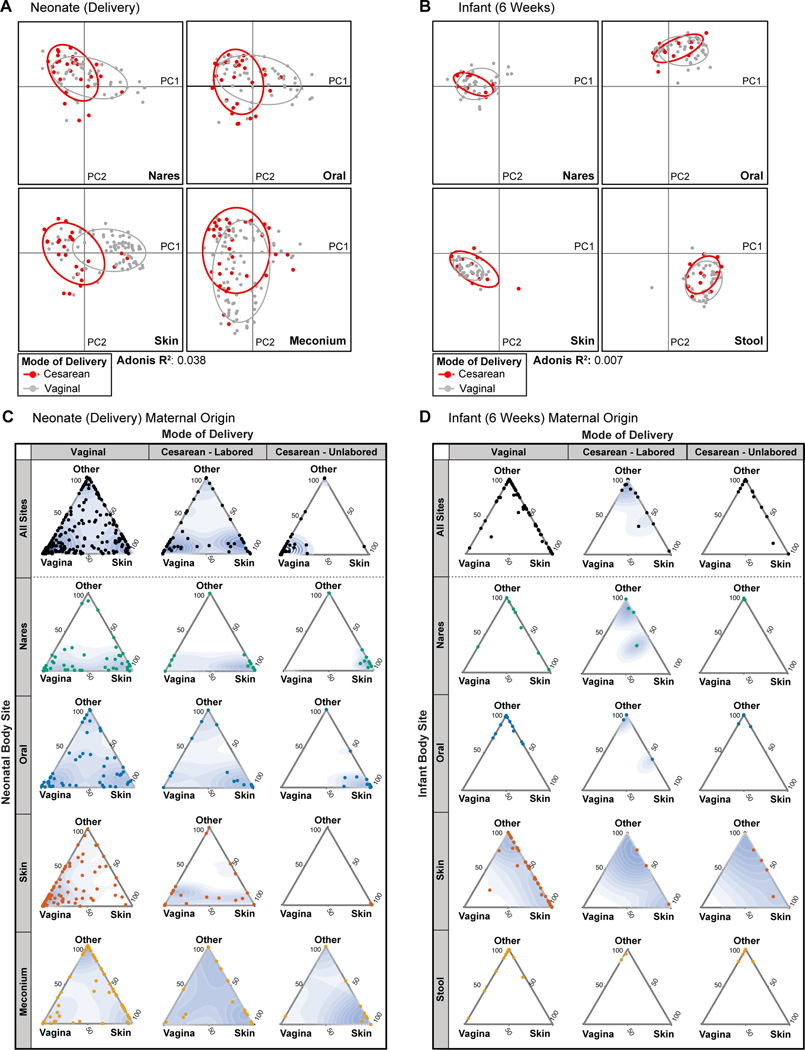
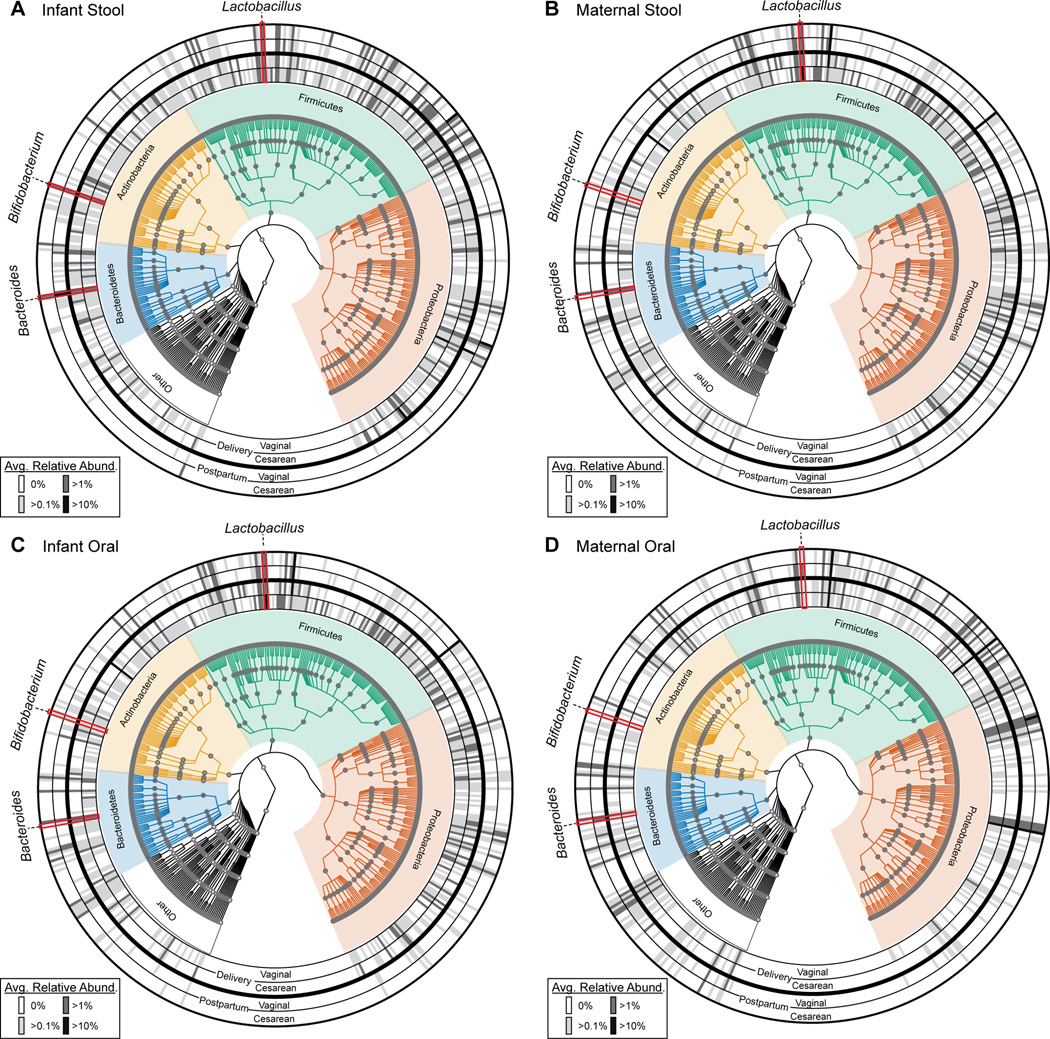
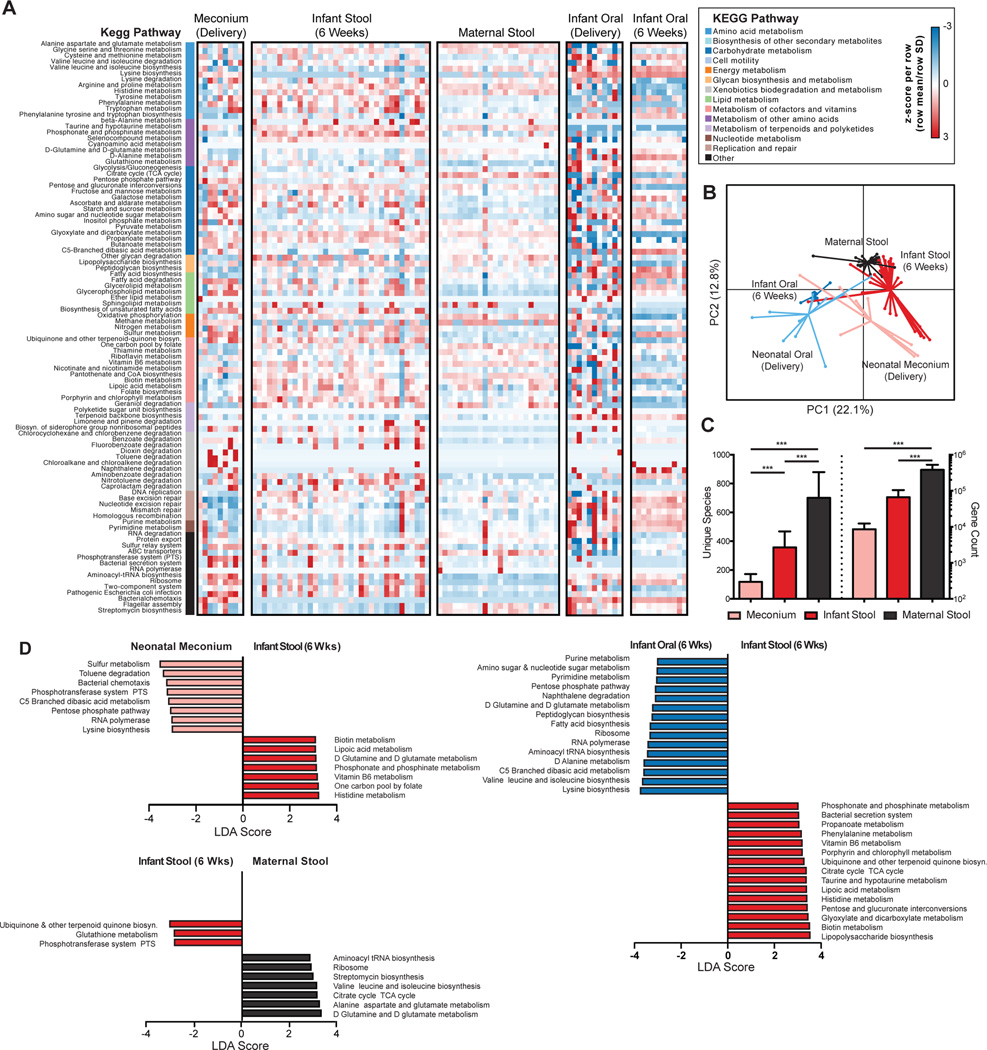
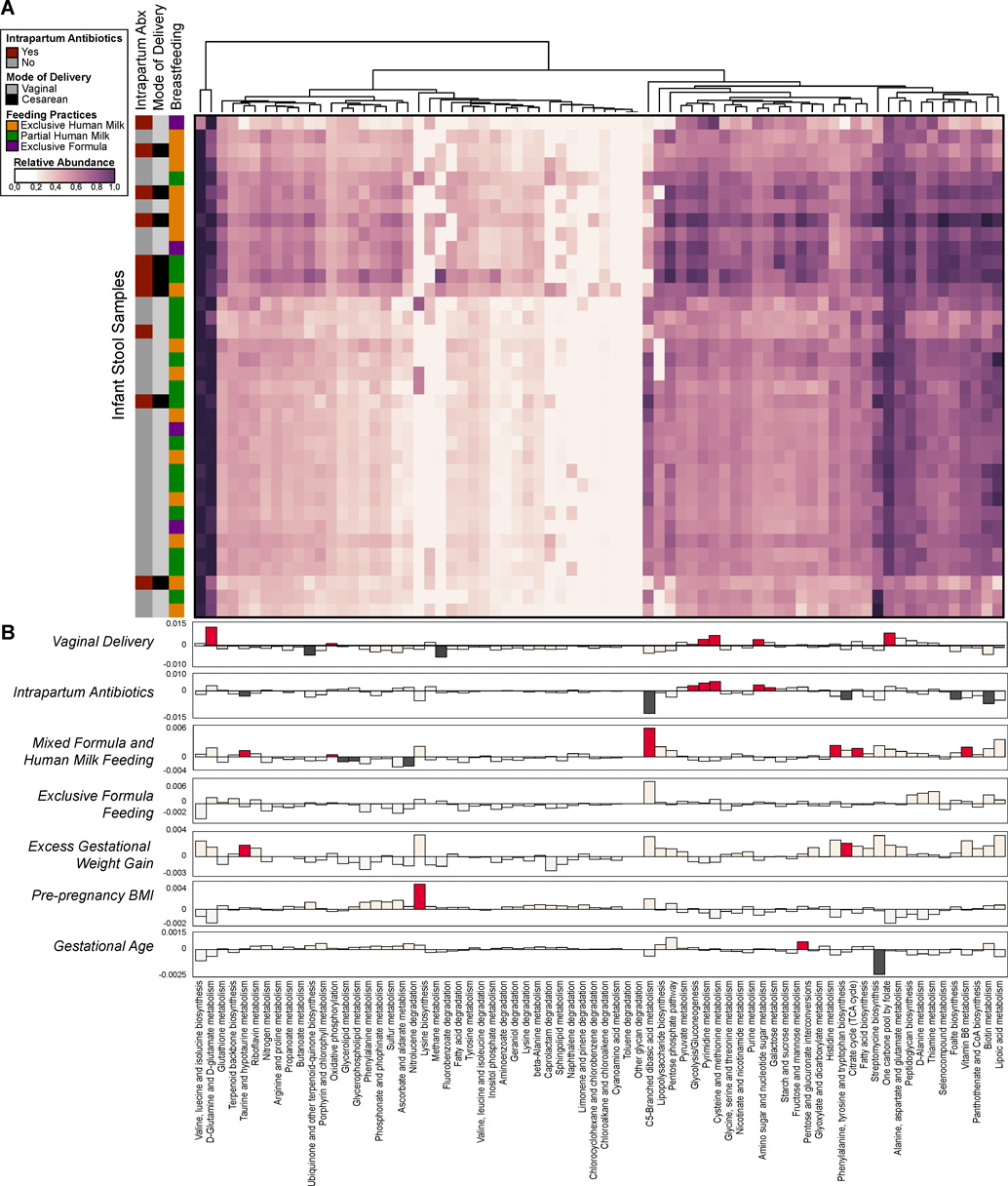
Human microbial communities are characterized by their taxonomic, metagenomic and metabolic diversity, which varies by distinct body sites and influences human physiology. However, when and how microbial communities within each body niche acquire unique taxonomical and functional signatures in early life remains underexplored. We thus sought to determine the taxonomic composition and potential metabolic function of the neonatal and early infant microbiota across multiple body sites and assess the effect of the mode of delivery and its potential confounders or modifiers. A cohort of pregnant women in their early third trimester (n = 81) were prospectively enrolled for longitudinal sampling through 6 weeks after delivery, and a second matched cross-sectional cohort (n = 81) was additionally recruited for sampling once at the time of delivery. Samples across multiple body sites, including stool, oral gingiva, nares, skin and vagina were collected for each maternal-infant dyad. Whole-genome shotgun sequencing and sequencing analysis of the gene encoding the 16S rRNA were performed to interrogate the composition and function of the neonatal and maternal microbiota. We found that the neonatal microbiota and its associated functional pathways were relatively homogeneous across all body sites at delivery, with the notable exception of the neonatal meconium. However, by 6 weeks after delivery, the infant microbiota structure and function had substantially expanded and diversified, with the body site serving as the primary determinant of the composition of the bacterial community and its functional capacity. Although minor variations in the neonatal (immediately at birth) microbiota community structure were associated with the cesarean mode of delivery in some body sites (oral gingiva, nares and skin; R2 = 0.038), this was not true for neonatal stool (meconium; Mann-Whitney P > 0.05), and there was no observable difference in community function regardless of delivery mode. For infants at 6 weeks of age, the microbiota structure and function had expanded and diversified with demonstrable body site specificity (P < 0.001, R2 = 0.189) but without discernable differences in community structure or function between infants delivered vaginally or by cesarean surgery (P = 0.057, R2 = 0.007). We conclude that within the first 6 weeks of life, the infant microbiota undergoes substantial reorganization, which is primarily driven by body site and not by mode of delivery.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Comment in
-
The shape of the microbiome in early life.Nat Med. 2017 Mar 7;23(3):274-275. doi: 10.1038/nm.4299. Nat Med. 2017. PMID: 28267714 No abstract available.
-
'Vaginal seeding' after a caesarean section provides benefits to newborn children: AGAINST: Vaginal microbiome transfer - a medical procedure with clear risks and uncertain benefits.BJOG. 2020 Jan;127(2):302. doi: 10.1111/1471-0528.15977. Epub 2019 Nov 6. BJOG. 2020. PMID: 31692198 No abstract available.
References
Main text References
-
- Turnbaugh PJ, et al. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006;444:1027–131. - PubMed
Methods-only References
-
- Edgar RC. Search and clustering orders of magnitude faster than BLAST. Bioinforma. Oxf. Engl. 2010;26:2460–2461. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
