

An intra-amygdala circuit specifically regulates social fear learning
- PMID: 28114293
- PMCID: PMC5323274
- DOI: 10.1038/nn.4481
An intra-amygdala circuit specifically regulates social fear learning
Abstract
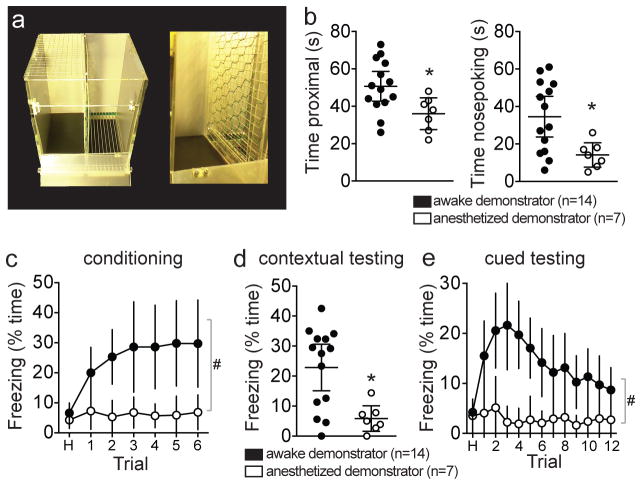
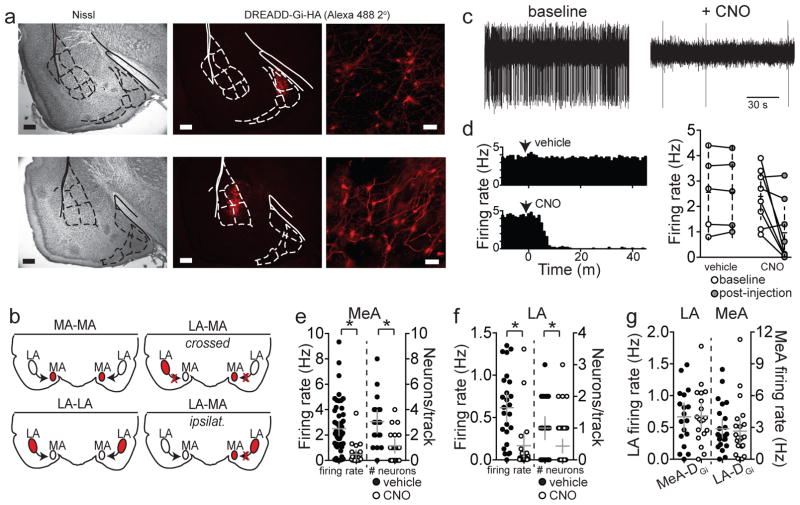
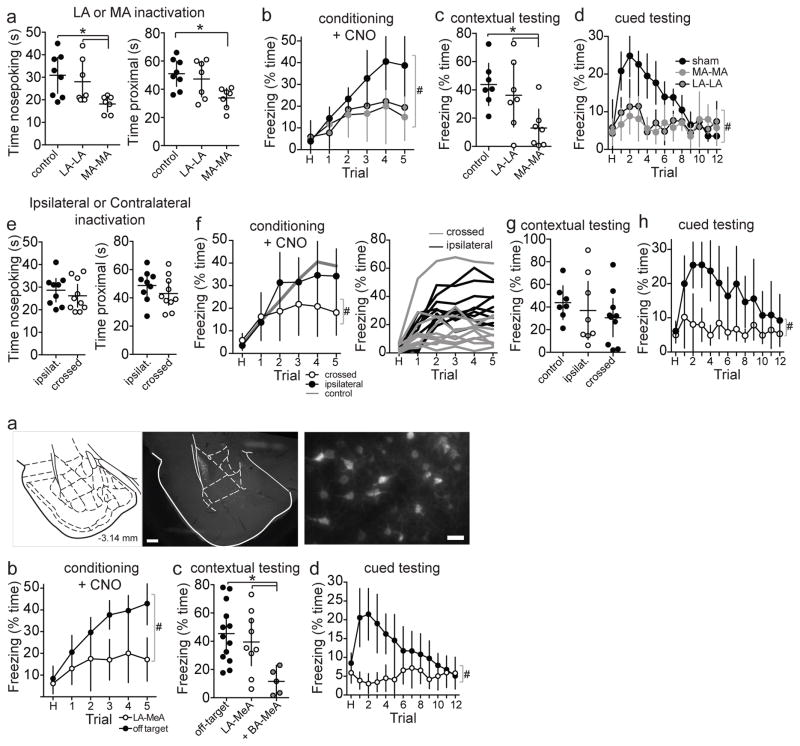
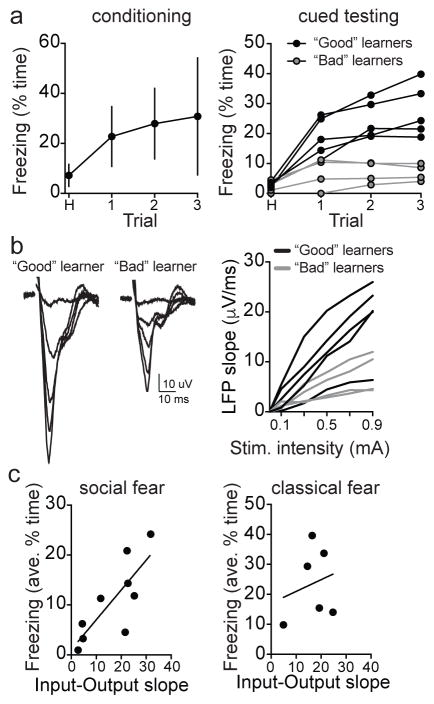
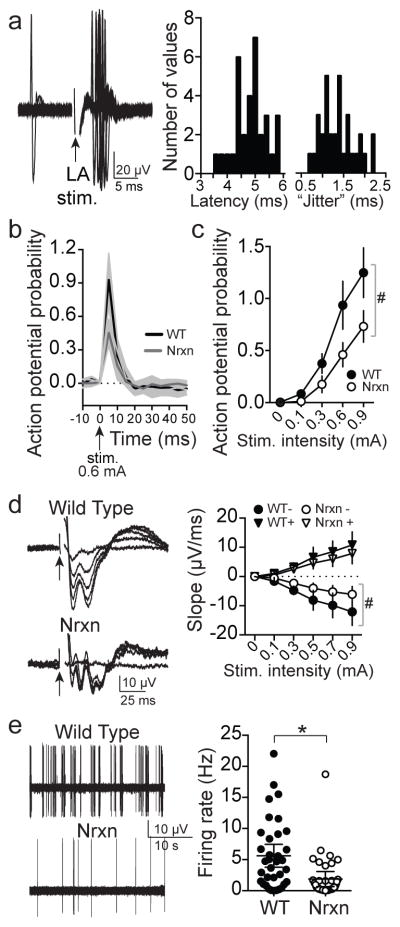
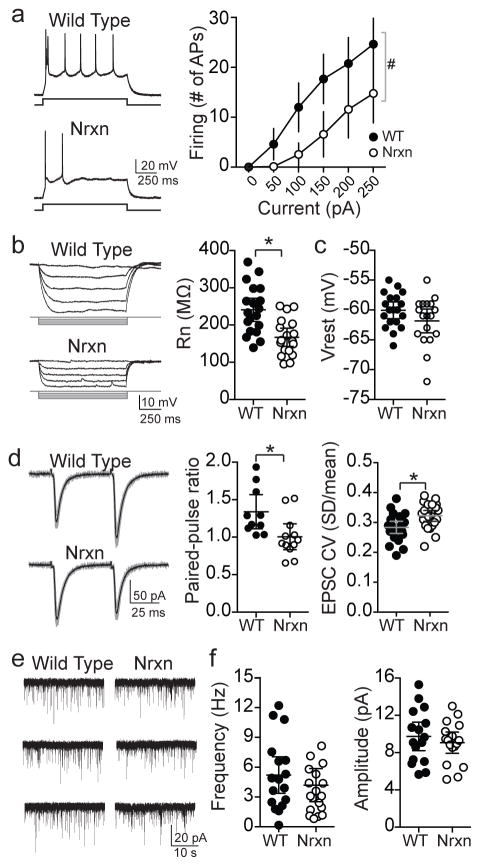
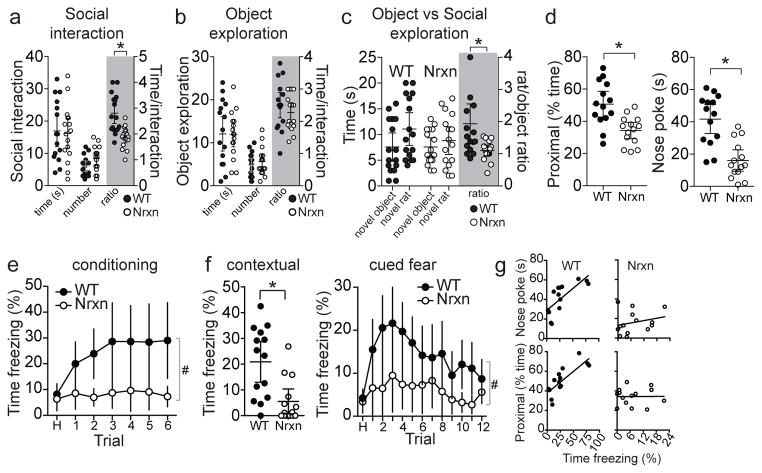
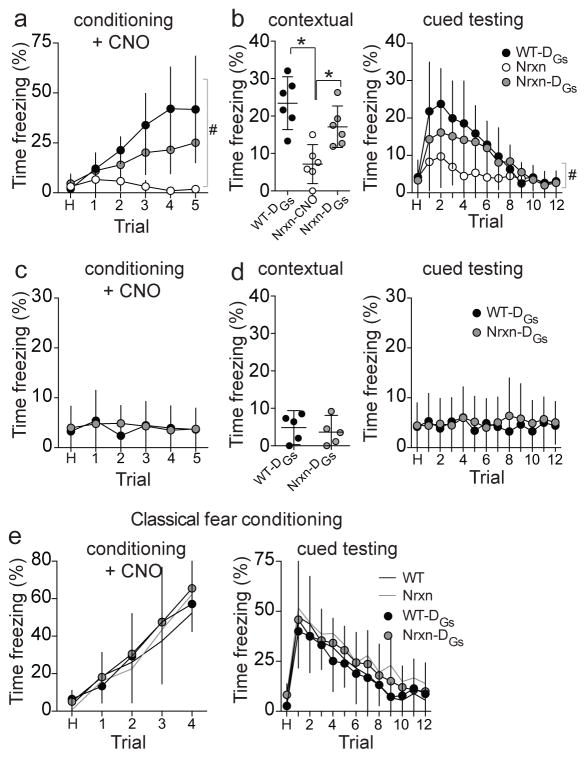
Adaptive social behavior requires transmission and reception of salient social information. Impairment of this reciprocity is a cardinal symptom of autism. The amygdala is a critical mediator of social behavior and is implicated in social symptoms of autism. Here we found that a specific amygdala circuit, from the lateral nucleus to the medial nucleus (LA-MeA), is required for using social cues to learn about environmental cues that signal imminent threats. Disruption of the LA-MeA circuit impaired valuation of these environmental cues and subsequent ability to use a cue to guide behavior. Rats with impaired social guidance of behavior due to knockout of Nrxn1, an analog of autism-associated gene NRXN, exhibited marked LA-MeA deficits. Chemogenetic activation of this circuit reversed these impaired social behaviors. These findings identify an amygdala circuit required to guide emotional responses to socially significant cues and identify an exploratory target for disorders associated with social impairments.
Conflict of interest statement
The authors report no competing financial interests.
Figures
Comment in
-
Rodent see, rodent fear.Nat Neurosci. 2017 Feb 23;20(3):381-382. doi: 10.1038/nn.4512. Nat Neurosci. 2017. PMID: 28230840 No abstract available.
References
-
- Adolphs R, Tranel D, Damasio H, Damasio A. Impaired recognition of emotion in facial expressions following bilateral damage to the human amygdala. Nature. 1994;372:669–672. - PubMed
-
- Brothers L, Ring B, Kling A. Response of neurons in the macaque amygdala to complex social stimuli. Behav Brain Res. 1990;41:199–213. - PubMed
-
- Dicks D, Myers RE, Kling A. Uncus and amiygdala lesions: effects on social behavior in the free-ranging rhesus monkey. Science. 1969;165:69–71. - PubMed
-
- Morris JS, et al. A differential neural response in the human amygdala to fearful and happy facial expressions. Nature. 1996;383:812–815. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
