

Morphological Transformation and Force Generation of Active Cytoskeletal Networks
- PMID: 28114384
- PMCID: PMC5256887
- DOI: 10.1371/journal.pcbi.1005277
Morphological Transformation and Force Generation of Active Cytoskeletal Networks
Abstract
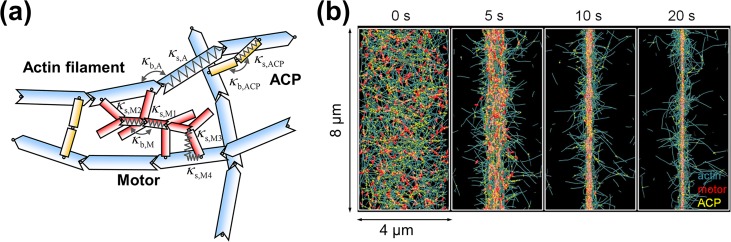
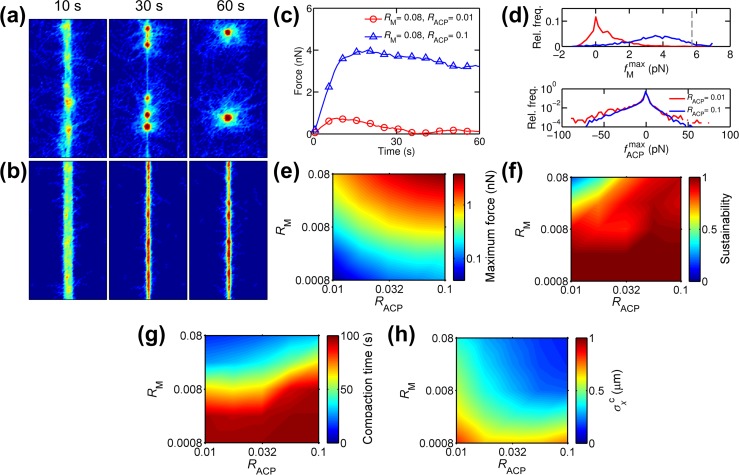
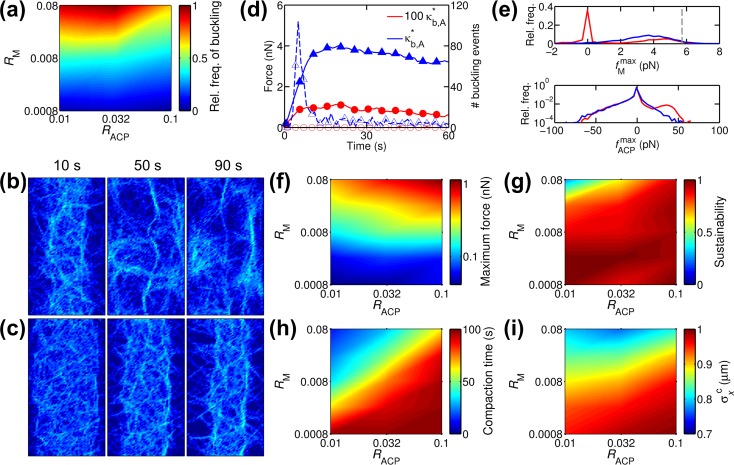
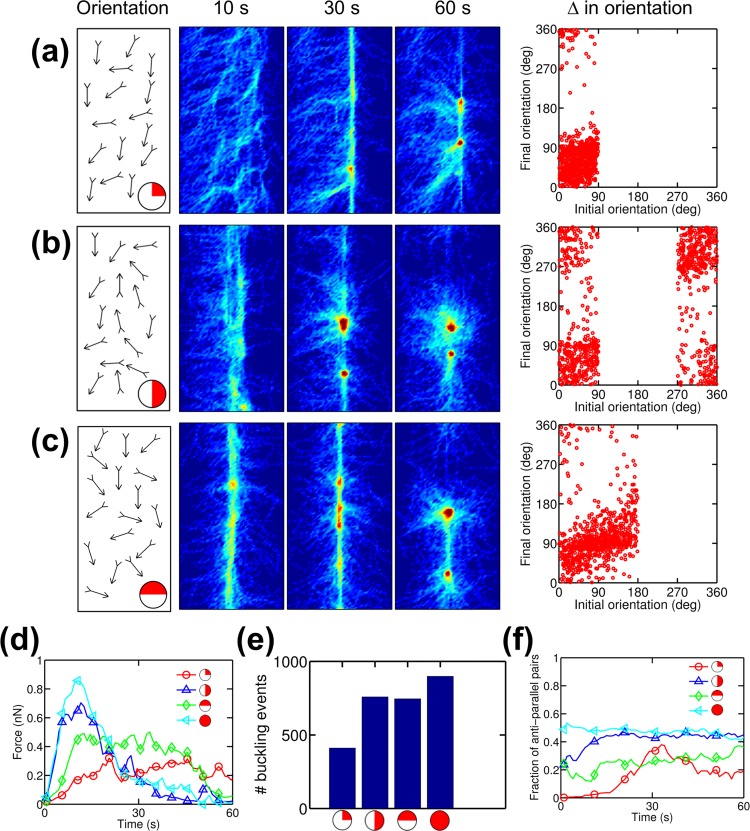
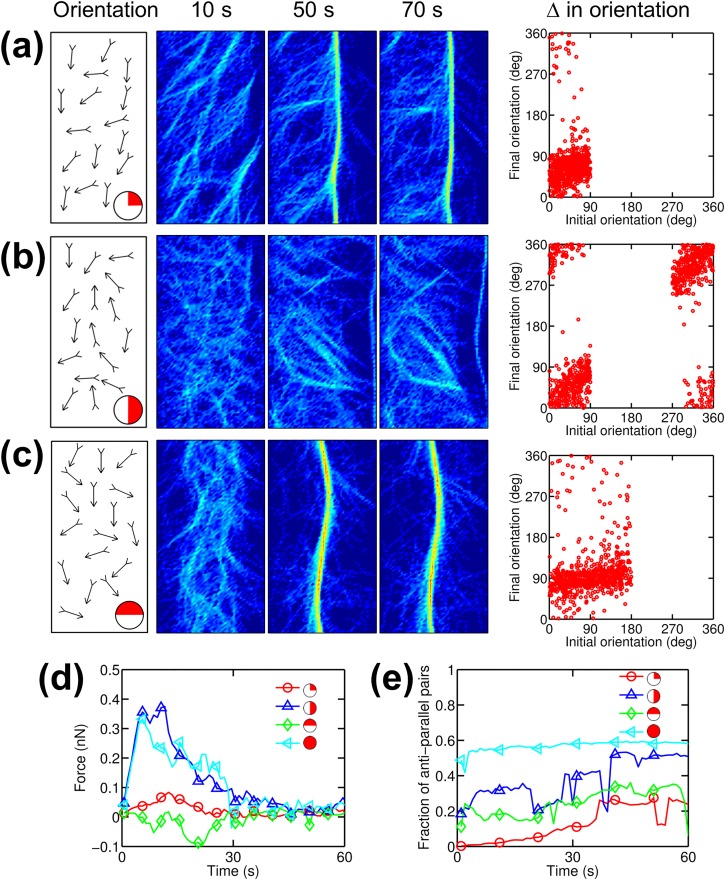
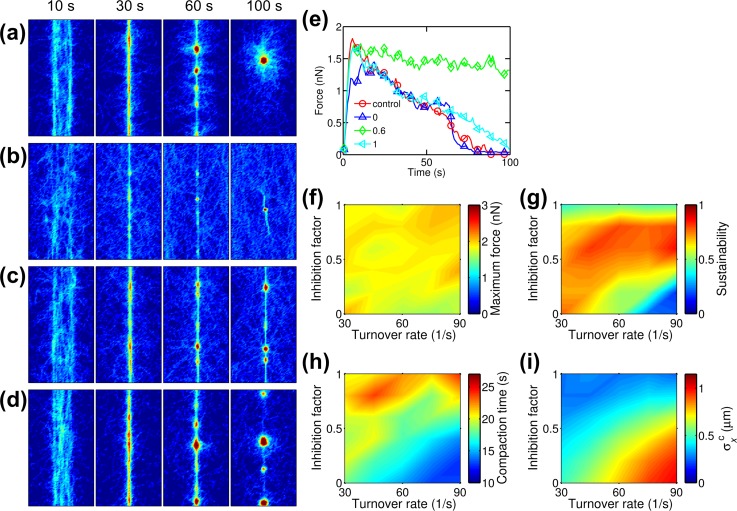
Cells assemble numerous types of actomyosin bundles that generate contractile forces for biological processes, such as cytokinesis and cell migration. One example of contractile bundles is a transverse arc that forms via actomyosin-driven condensation of actin filaments in the lamellipodia of migrating cells and exerts significant forces on the surrounding environments. Structural reorganization of a network into a bundle facilitated by actomyosin contractility is a physiologically relevant and biophysically interesting process. Nevertheless, it remains elusive how actin filaments are reoriented, buckled, and bundled as well as undergo tension buildup during the structural reorganization. In this study, using an agent-based computational model, we demonstrated how the interplay between the density of myosin motors and cross-linking proteins and the rigidity, initial orientation, and turnover of actin filaments regulates the morphological transformation of a cross-linked actomyosin network into a bundle and the buildup of tension occurring during the transformation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
