

Subinhibitory Concentrations of Bacteriostatic Antibiotics Induce relA-Dependent and relA-Independent Tolerance to β-Lactams
- PMID: 28115345
- PMCID: PMC5365698
- DOI: 10.1128/AAC.02173-16
Subinhibitory Concentrations of Bacteriostatic Antibiotics Induce relA-Dependent and relA-Independent Tolerance to β-Lactams
Abstract
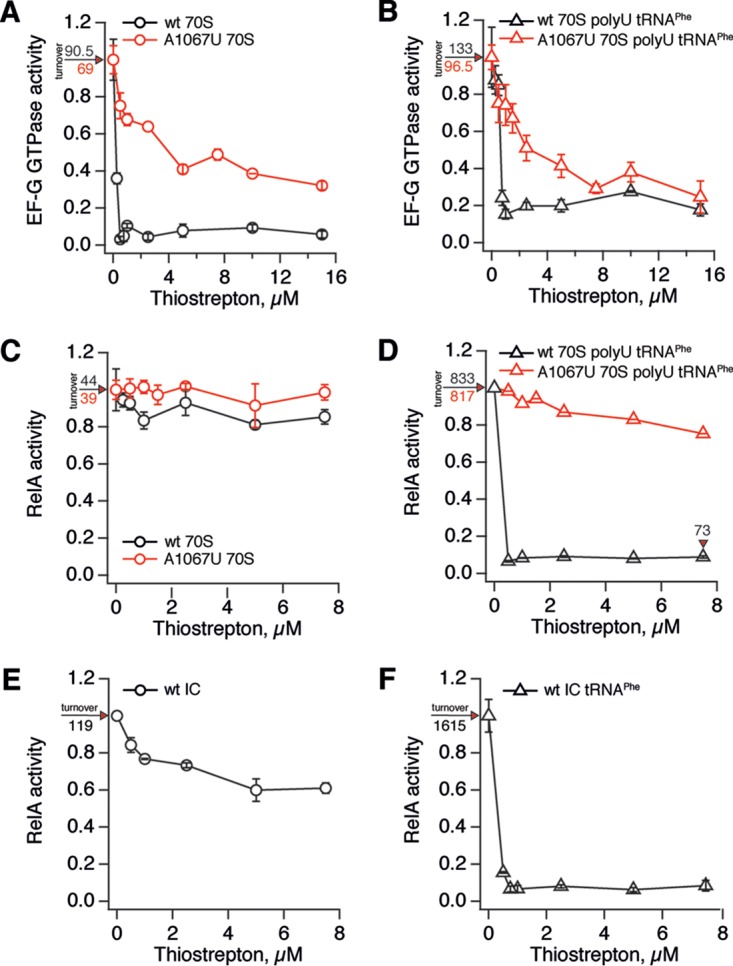
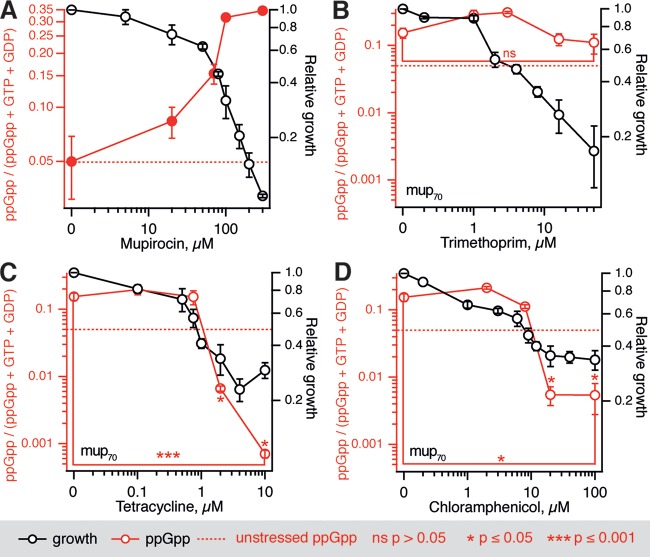
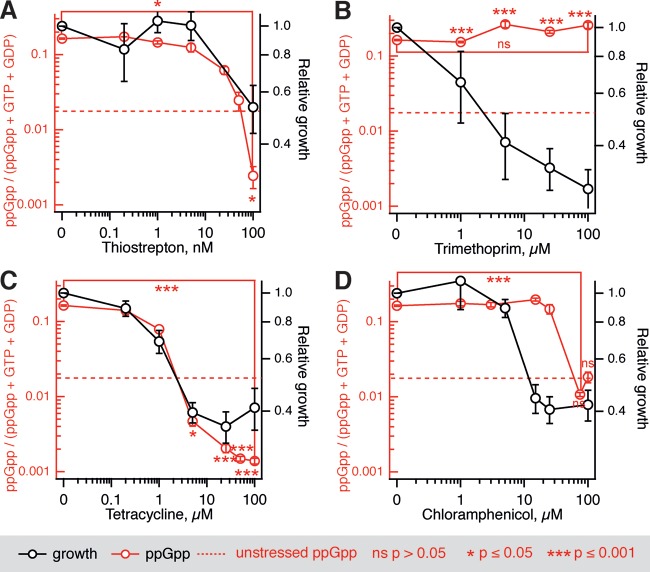
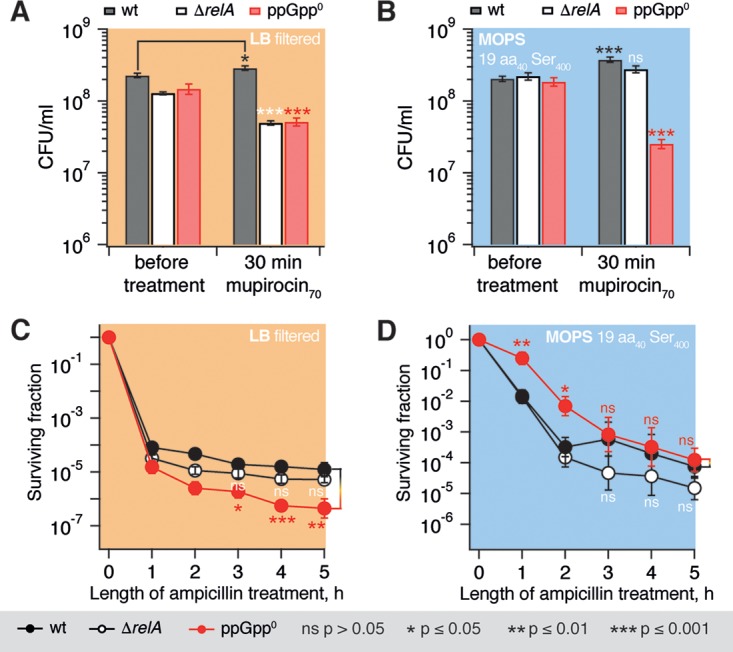
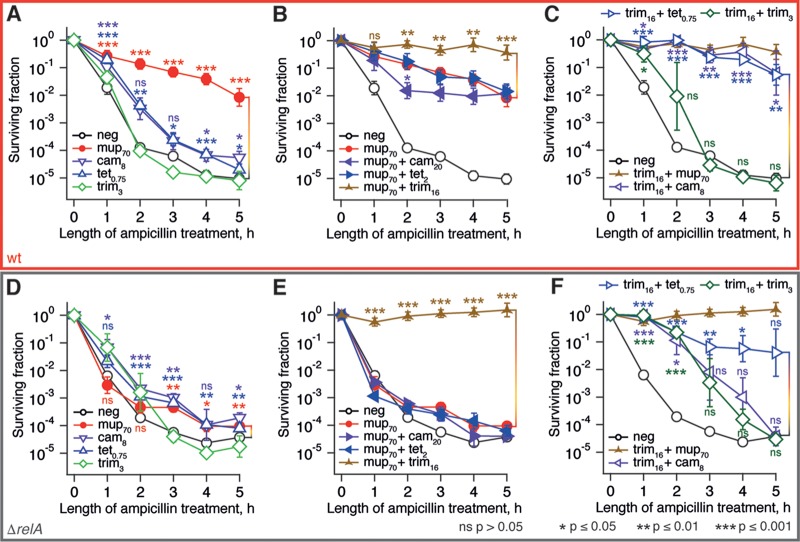
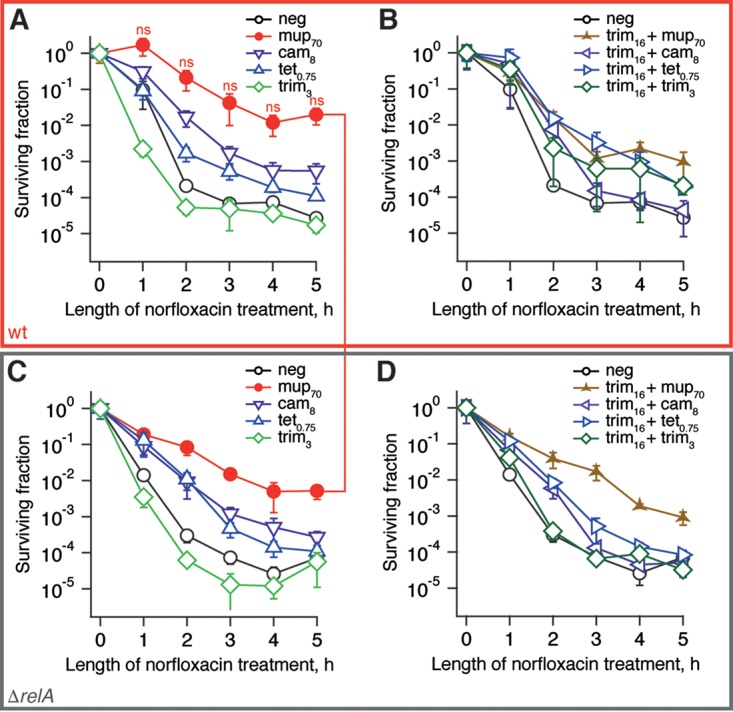
The nucleotide (p)ppGpp is a key regulator of bacterial metabolism, growth, stress tolerance, and virulence. During amino acid starvation, the Escherichia coli (p)ppGpp synthetase RelA is activated by deacylated tRNA in the ribosomal A-site. An increase in (p)ppGpp is believed to drive the formation of antibiotic-tolerant persister cells, prompting the development of strategies to inhibit (p)ppGpp synthesis. We show that in a biochemical system from purified E. coli components, the antibiotic thiostrepton efficiently inhibits RelA activation by the A-site tRNA. In bacterial cultures, the ribosomal inhibitors thiostrepton, chloramphenicol, and tetracycline all efficiently abolish accumulation of (p)ppGpp induced by the Ile-tRNA synthetase inhibitor mupirocin. This abolishment, however, does not reduce the persister level. In contrast, the combination of dihydrofolate reductase inhibitor trimethoprim with mupirocin, tetracycline, or chloramphenicol leads to ampicillin tolerance. The effect is independent of RelA functionality, specific to β-lactams, and not observed with the fluoroquinolone norfloxacin. These results refine our understanding of (p)ppGpp's role in antibiotic tolerance and persistence and demonstrate unexpected drug interactions that lead to tolerance to bactericidal antibiotics.
Keywords: RelA; antibiotics; mupirocin; persistence; ppGpp; ribosomes; thiostrepton; tolerance; trimethoprim; β-lactam.
Copyright © 2017 Kudrin et al.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
