

The IQD Family of Calmodulin-Binding Proteins Links Calcium Signaling to Microtubules, Membrane Subdomains, and the Nucleus
- PMID: 28115582
- PMCID: PMC5338658
- DOI: 10.1104/pp.16.01743
The IQD Family of Calmodulin-Binding Proteins Links Calcium Signaling to Microtubules, Membrane Subdomains, and the Nucleus
Abstract
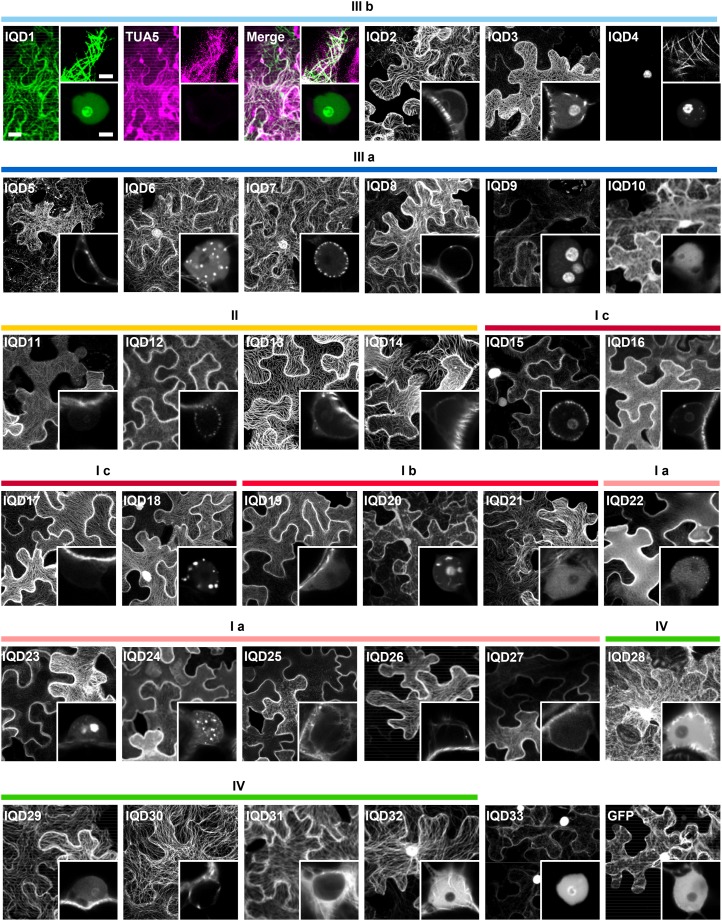
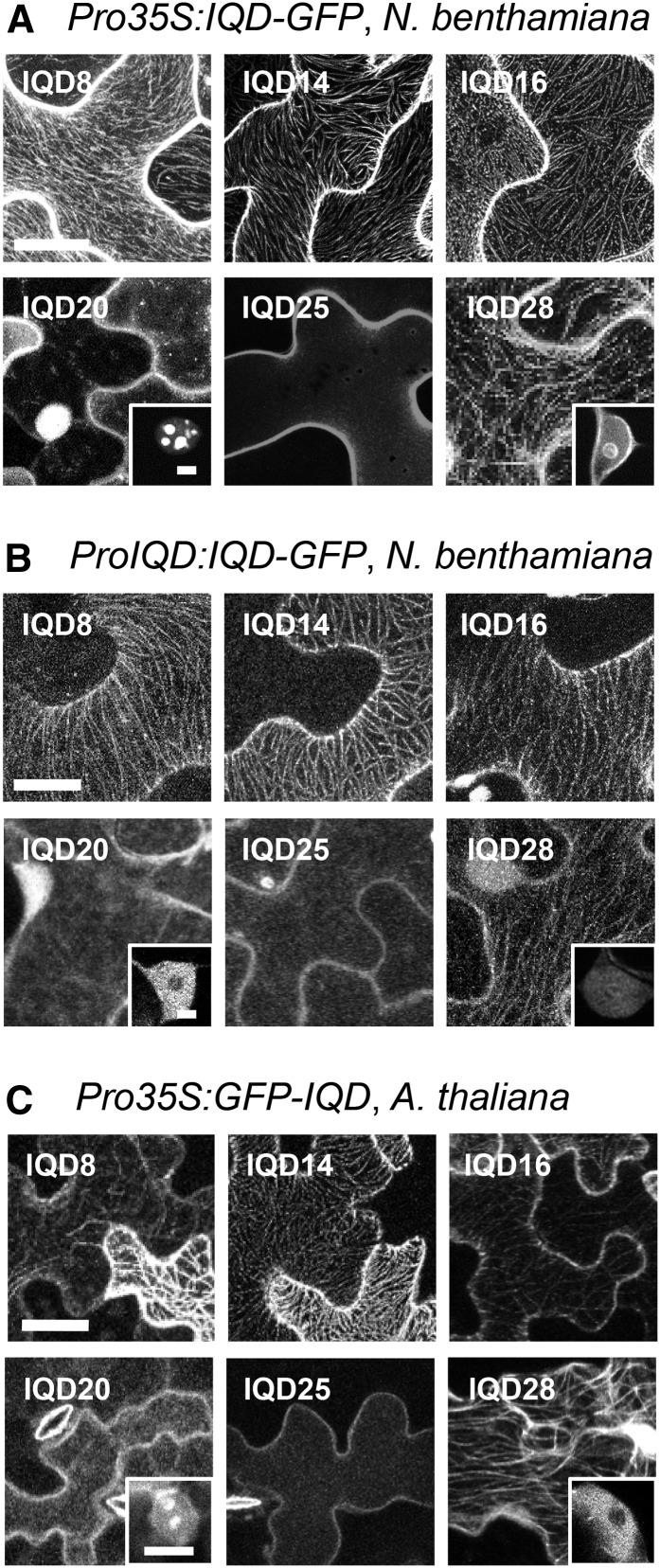
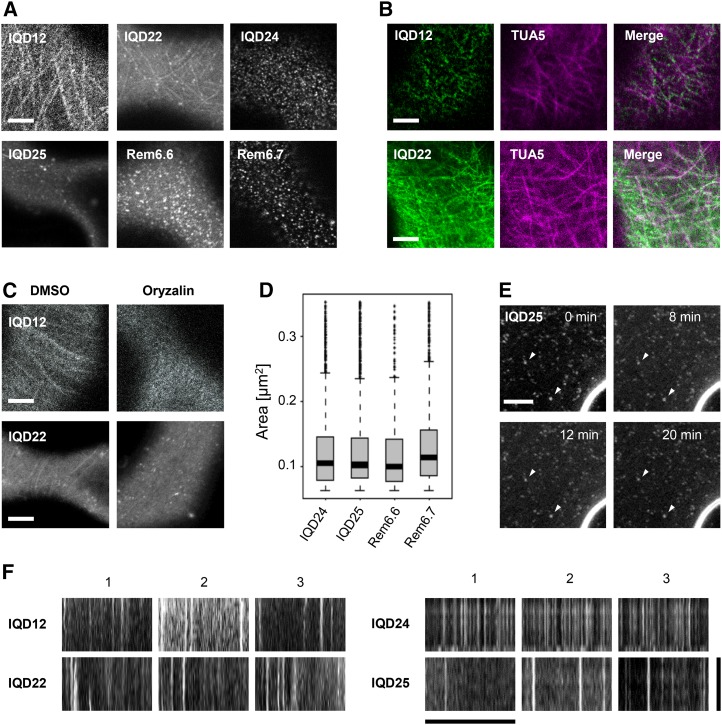
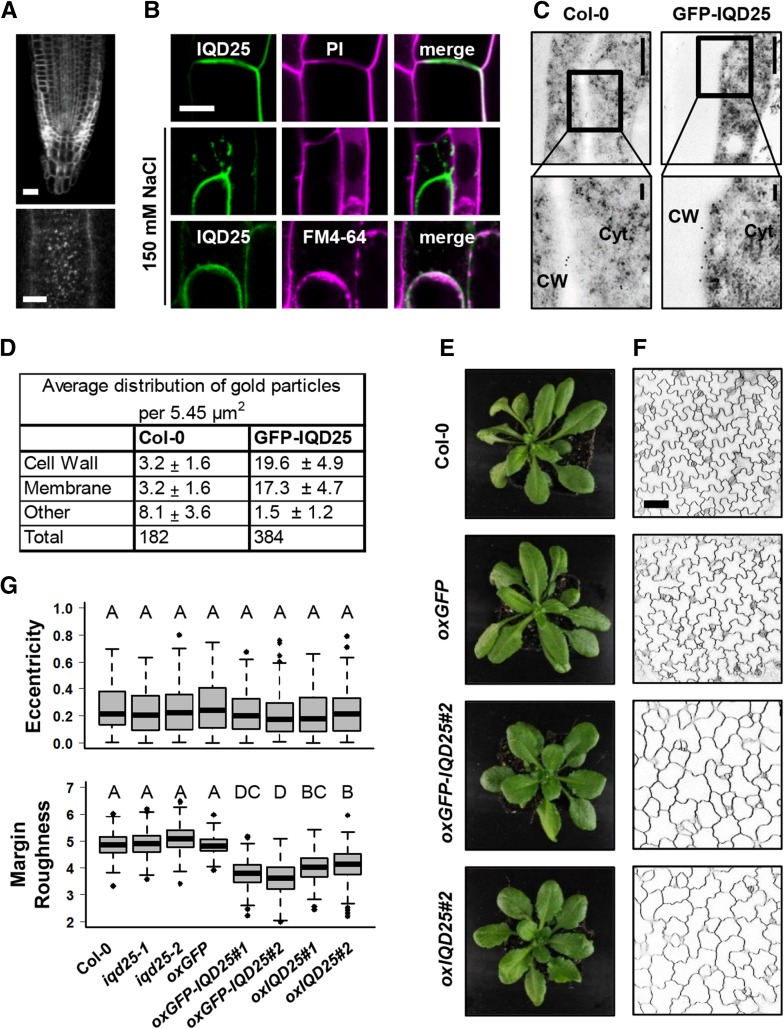
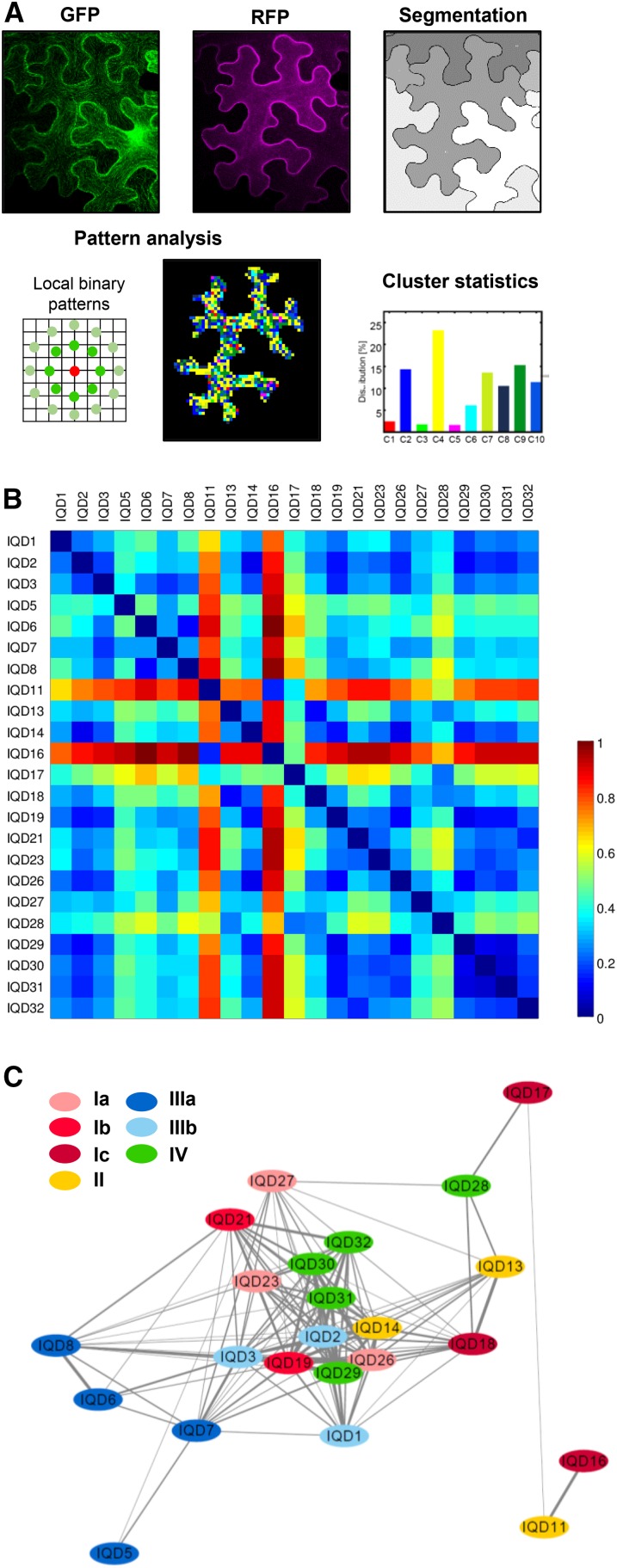
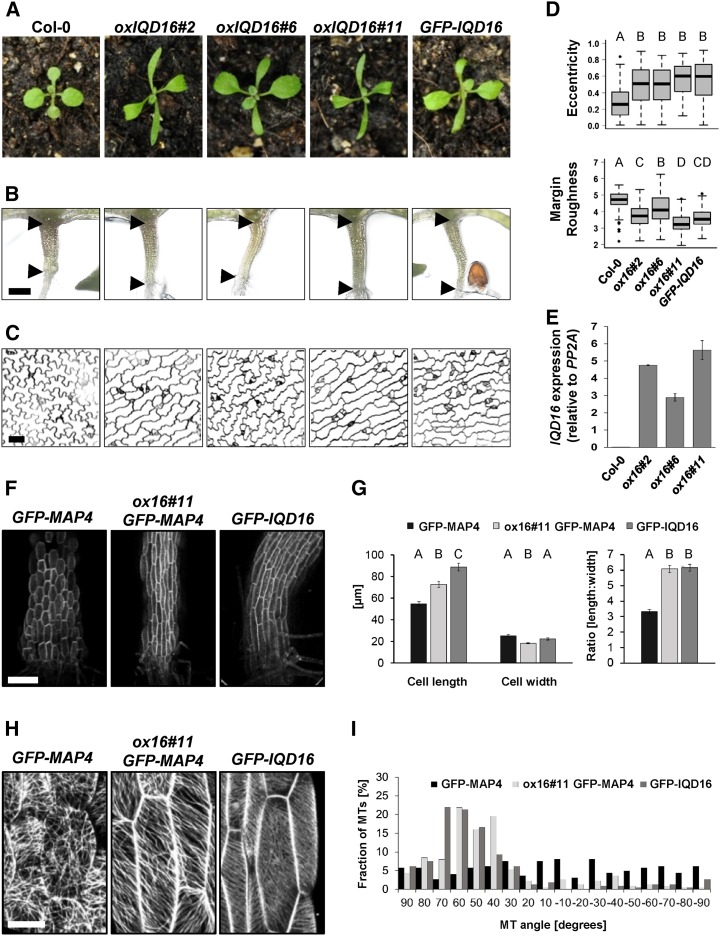
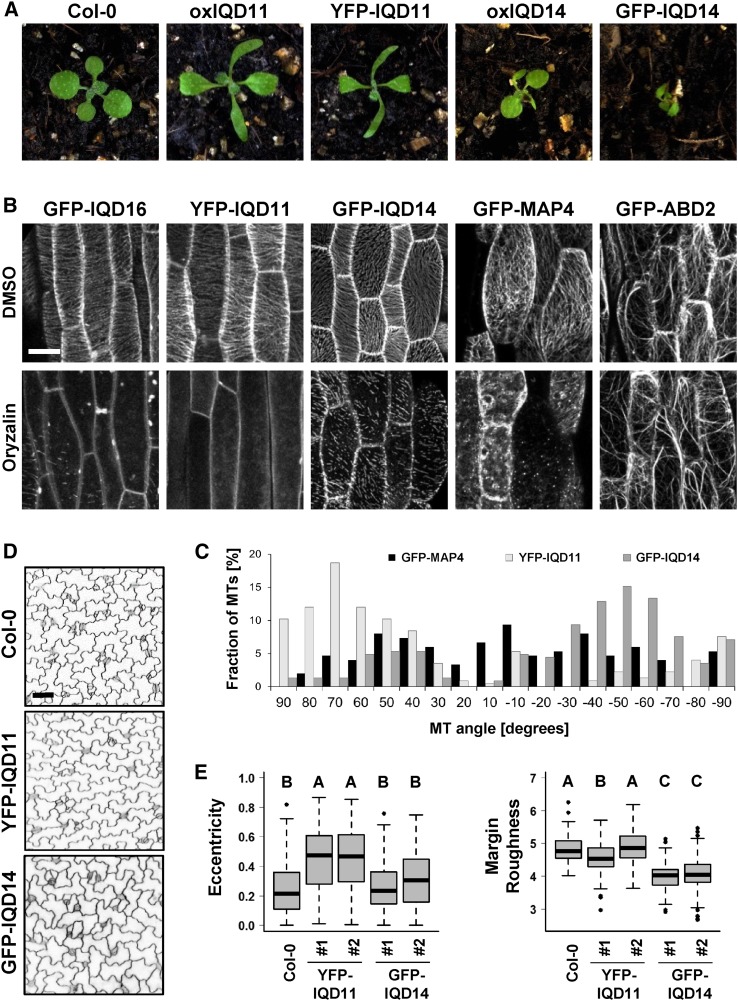
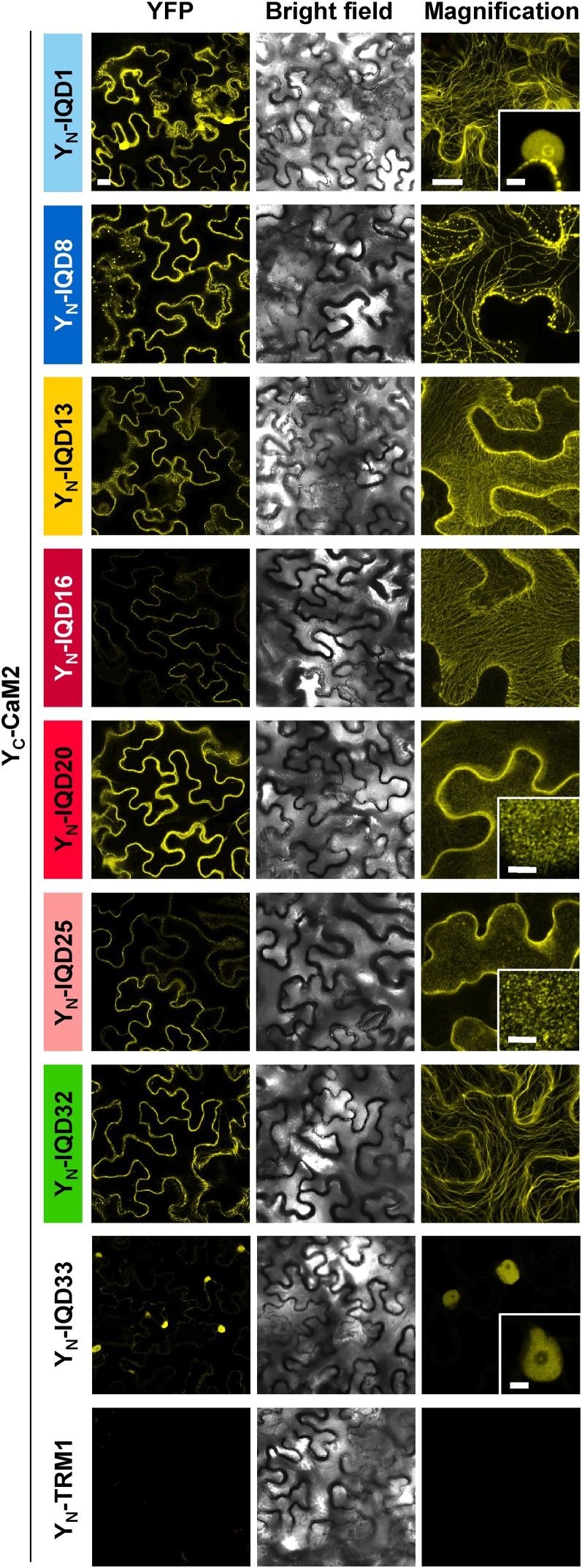
Calcium (Ca2+) signaling and dynamic reorganization of the cytoskeleton are essential processes for the coordination and control of plant cell shape and cell growth. Calmodulin (CaM) and closely related calmodulin-like (CML) polypeptides are principal sensors of Ca2+ signals. CaM/CMLs decode and relay information encrypted by the second messenger via differential interactions with a wide spectrum of targets to modulate their diverse biochemical activities. The plant-specific IQ67 DOMAIN (IQD) family emerged as possibly the largest class of CaM-interacting proteins with undefined molecular functions and biological roles. Here, we show that the 33 members of the IQD family in Arabidopsis (Arabidopsis thaliana) differentially localize, using green fluorescent protein (GFP)-tagged proteins, to multiple and distinct subcellular sites, including microtubule (MT) arrays, plasma membrane subdomains, and nuclear compartments. Intriguingly, the various IQD-specific localization patterns coincide with the subcellular patterns of IQD-dependent recruitment of CaM, suggesting that the diverse IQD members sequester Ca2+-CaM signaling modules to specific subcellular sites for precise regulation of Ca2+-dependent processes. Because MT localization is a hallmark of most IQD family members, we quantitatively analyzed GFP-labeled MT arrays in Nicotiana benthamiana cells transiently expressing GFP-IQD fusions and observed IQD-specific MT patterns, which point to a role of IQDs in MT organization and dynamics. Indeed, stable overexpression of select IQD proteins in Arabidopsis altered cellular MT orientation, cell shape, and organ morphology. Because IQDs share biochemical properties with scaffold proteins, we propose that IQD families provide an assortment of platform proteins for integrating CaM-dependent Ca2+ signaling at multiple cellular sites to regulate cell function, shape, and growth.
© 2017 American Society of Plant Biologists. All Rights Reserved.
Figures
Comment in
-
On the Inside.Plant Physiol. 2017 Mar;173(3):1527-1528. doi: 10.1104/pp.17.00218. Plant Physiol. 2017. PMID: 28258119 Free PMC article. No abstract available.
References
-
- Babu MM, van der Lee R, de Groot NS, Gsponer J (2011) Intrinsically disordered proteins: regulation and disease. Curr Opin Struct Biol 21: 432–440 - PubMed
-
- Benschop JJ, Mohammed S, O’Flaherty M, Heck AJR, Slijper M, Menke FLH (2007) Quantitative phosphoproteomics of early elicitor signaling in Arabidopsis. Mol Cell Proteomics 6: 1198–1214 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
