

Durum Wheat Roots Adapt to Salinity Remodeling the Cellular Content of Nitrogen Metabolites and Sucrose
- PMID: 28119716
- PMCID: PMC5220018
- DOI: 10.3389/fpls.2016.02035
Durum Wheat Roots Adapt to Salinity Remodeling the Cellular Content of Nitrogen Metabolites and Sucrose
Abstract
Plants are currently experiencing increasing salinity problems due to irrigation with brackish water. Moreover, in fields, roots can grow in soils which show spatial variation in water content and salt concentration, also because of the type of irrigation. Salinity impairs crop growth and productivity by inhibiting many physiological and metabolic processes, in particular nitrate uptake, translocation, and assimilation. Salinity determines an increase of sap osmolality from about 305 mOsmol kg-1 in control roots to about 530 mOsmol kg-1 in roots under salinity. Root cells adapt to salinity by sequestering sodium in the vacuole, as a cheap osmoticum, and showing a rearrangement of few nitrogen-containing metabolites and sucrose in the cytosol, both for osmotic adjustment and oxidative stress protection, thus providing plant viability even at low nitrate levels. Mainly glycine betaine and sucrose at low nitrate concentration, and glycine betaine, asparagine and proline at high nitrate levels can be assumed responsible for the osmotic adjustment of the cytosol, the assimilation of the excess of ammonium and the scavenging of ROS under salinity. High nitrate plants with half of the root system under salinity accumulate proline and glutamine in both control and salt stressed split roots, revealing that osmotic adjustment is not a regional effect in plants. The expression level and enzymatic activities of asparagine synthetase and Δ1-pyrroline-5-carboxylate synthetase, as well as other enzymatic activities of nitrogen and carbon metabolism, are analyzed.
Keywords: P5CS; asparagine; asparagine synthetase; glycine betaine; nitrate reductase; osmotic adjustment.
Figures
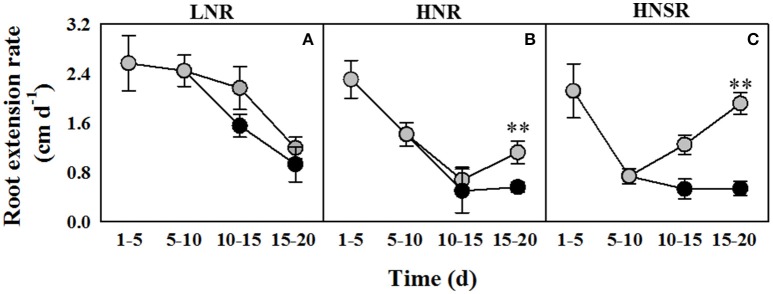
 ) or salt (100 mM NaCl,
) or salt (100 mM NaCl,  ) conditions. Six replicate plants of each treatment were measured on days 5, 10, 15, and 20 of hydroponic culture. was added on day 5 and 100 mM NaCl was added from day 10. The values are means ± SD (n = 6). Significant differences between treatments are indicated by asterisks (**p < 0.01; LSD-test).
) conditions. Six replicate plants of each treatment were measured on days 5, 10, 15, and 20 of hydroponic culture. was added on day 5 and 100 mM NaCl was added from day 10. The values are means ± SD (n = 6). Significant differences between treatments are indicated by asterisks (**p < 0.01; LSD-test).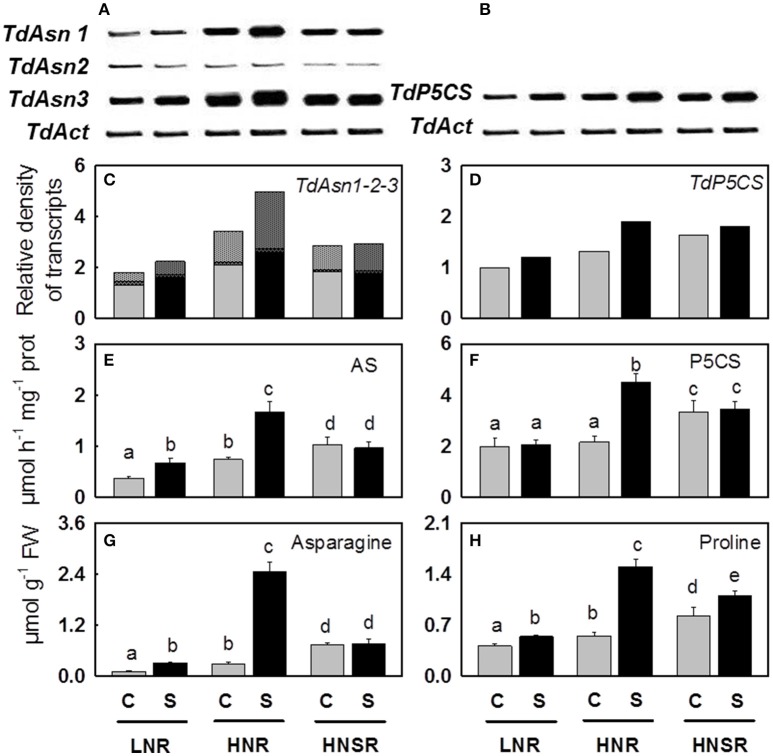
 ) and salt stressed (
) and salt stressed ( ) plants. Plants were subjected to salt stress starting from day 10 of culture. Control plants were grown without NaCl addition. Plants were harvested after 20 days of hydroponic culture. The values are mean ± SD (n = 4). Different letters above bars indicate significant difference between treatments (p < 0.05, LSD-test).
) plants. Plants were subjected to salt stress starting from day 10 of culture. Control plants were grown without NaCl addition. Plants were harvested after 20 days of hydroponic culture. The values are mean ± SD (n = 4). Different letters above bars indicate significant difference between treatments (p < 0.05, LSD-test).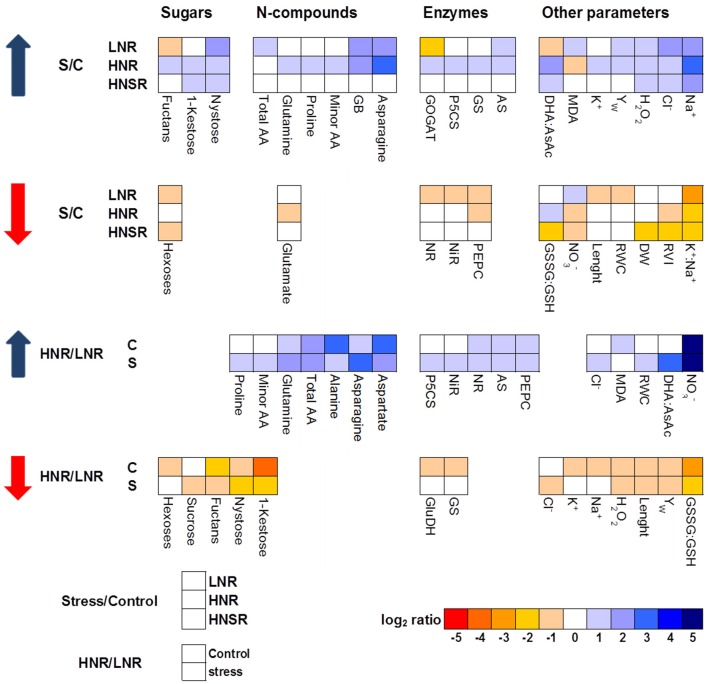
References
-
- Abd-El-Baki G. K., Siefritz F., Man H. M., Weiner H., Kaldenhoff R., Kaiser W. M. (2000). Nitrate reductase in Zea mays L. under salinity. Plant Cell Environ. 23, 515–521. 10.1046/j.1365-3040.2000.00568.x - DOI
-
- Annunziata M. G., Attico A., Woodrow P., Oliva M. A., Fuggi A., Carillo P. (2012). An improved fluorimetric HPLC method for quantifying tocopherols in Brassica rapa L. subsp. sylvestris after harvest. J. Food Composition Anal. 27, 145–150. 10.1016/j.jfca.2012.05.006 - DOI
-
- Ashraf M., Foolad M. R. (2007). Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 59, 206–216. 10.1016/j.envexpbot.2005.12.006 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous