

Culture and biology in the origins of linguistic structure
- PMID: 28120320
- PMCID: PMC5325872
- DOI: 10.3758/s13423-016-1166-7
Culture and biology in the origins of linguistic structure
Abstract
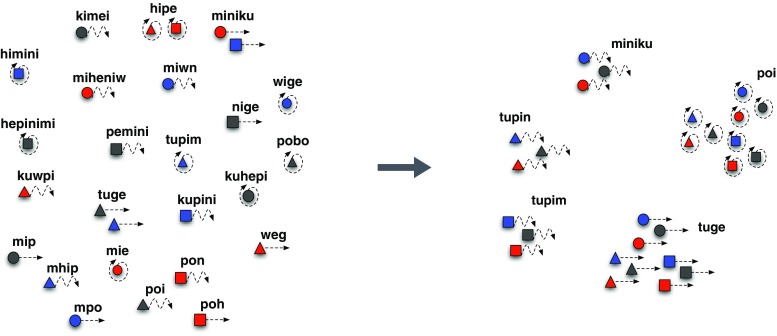
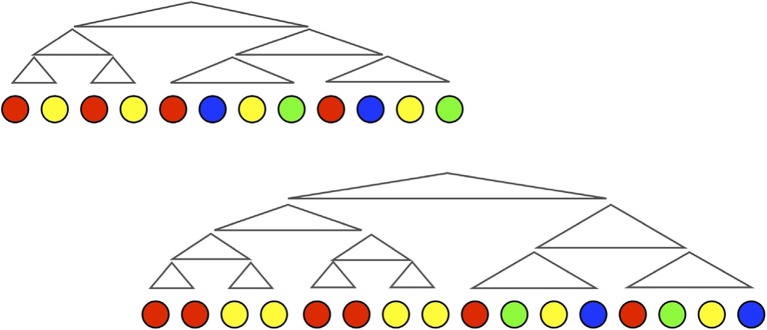
Language is systematically structured at all levels of description, arguably setting it apart from all other instances of communication in nature. In this article, I survey work over the last 20 years that emphasises the contributions of individual learning, cultural transmission, and biological evolution to explaining the structural design features of language. These 3 complex adaptive systems exist in a network of interactions: individual learning biases shape the dynamics of cultural evolution; universal features of linguistic structure arise from this cultural process and form the ultimate linguistic phenotype; the nature of this phenotype affects the fitness landscape for the biological evolution of the language faculty; and in turn this determines individuals' learning bias. Using a combination of computational simulation, laboratory experiments, and comparison with real-world cases of language emergence, I show that linguistic structure emerges as a natural outcome of cultural evolution once certain minimal biological requirements are in place.
Keywords: Computational modeling; Cultural evolution; Iterated learning; Language evolution.
Figures








Comment in
-
The continuing legacy of nature versus nurture in biolinguistics.Psychon Bull Rev. 2017 Feb;24(1):140-141. doi: 10.3758/s13423-016-1202-7. Psychon Bull Rev. 2017. PMID: 28120321 Free PMC article.
-
Restrictiveness matters.Psychon Bull Rev. 2017 Feb;24(1):138-139. doi: 10.3758/s13423-016-1194-3. Psychon Bull Rev. 2017. PMID: 28120322 Free PMC article.
References
-
- Andersen H. Abductive and deductive change. Language. 1973;49(4):765–793. doi: 10.2307/412063. - DOI
-
- Arbib MA, Bickerton D. The emergence of protolanguage: Holophrasis vs compositionality. Amsterdam, The Netherlands: John Benjamins; 2010.
-
- Bartlett FC. Remembering: An experimental and social study. Cambridge, UK: Cambridge University; 1932.
-
- Batali, J. (1998). Computational simulations of the emergence of grammar. In J. Hurford, M. Studdert-Kennedy & C. Knight (Eds.), Approaches to the Evolution of Language: Social and Cognitive Bases (pp. 405-426). Cambridge, UK: Cambridge University Press.
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
