

The Matrix protein M1 from influenza C virus induces tubular membrane invaginations in an in vitro cell membrane model
- PMID: 28120862
- PMCID: PMC5264427
- DOI: 10.1038/srep40801
The Matrix protein M1 from influenza C virus induces tubular membrane invaginations in an in vitro cell membrane model
Abstract
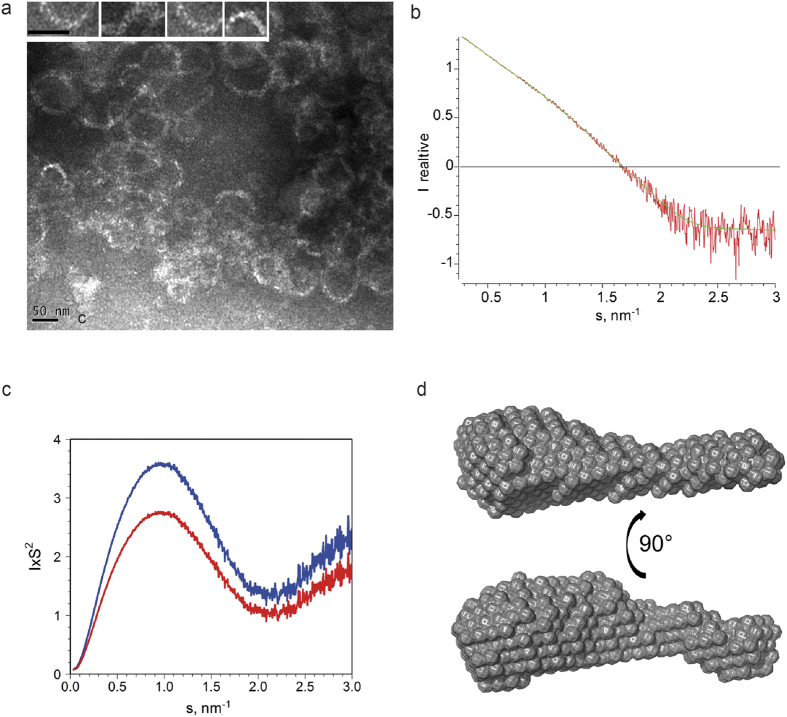
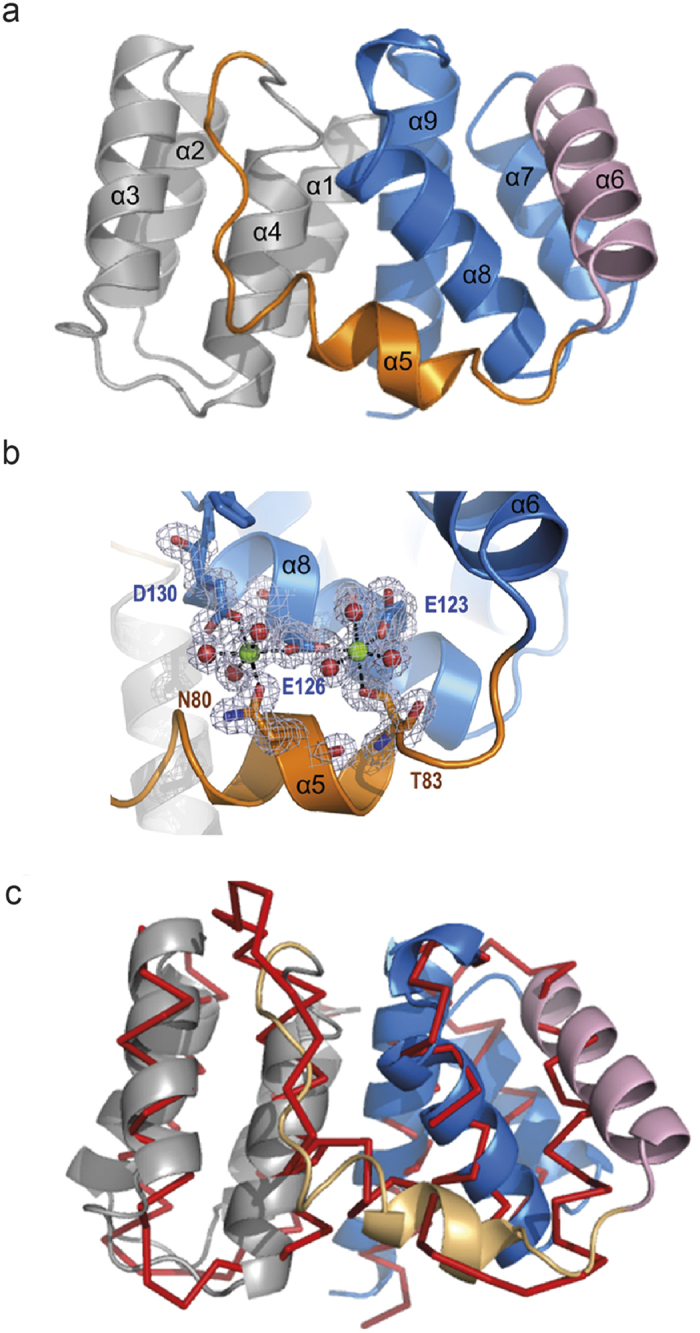
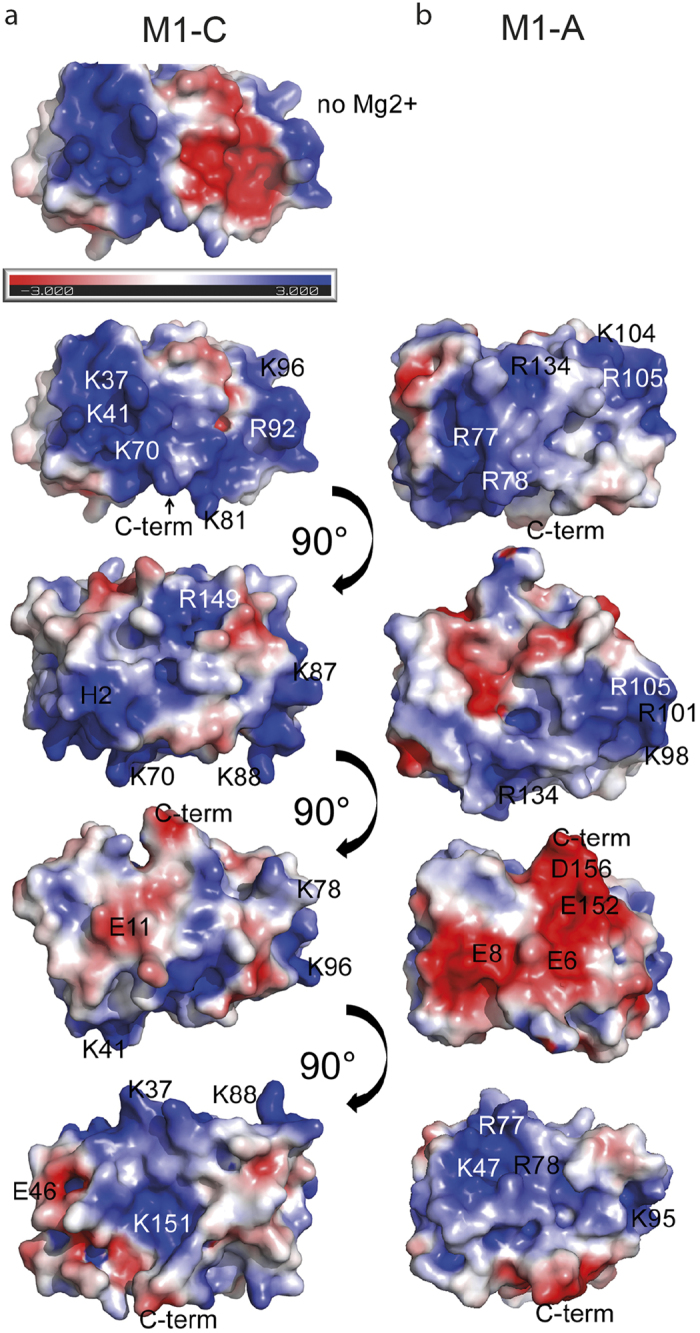
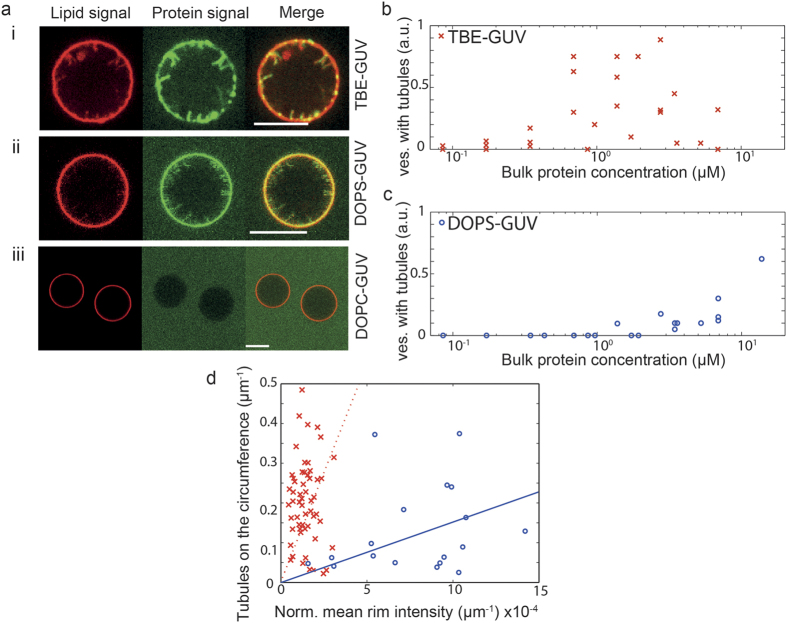
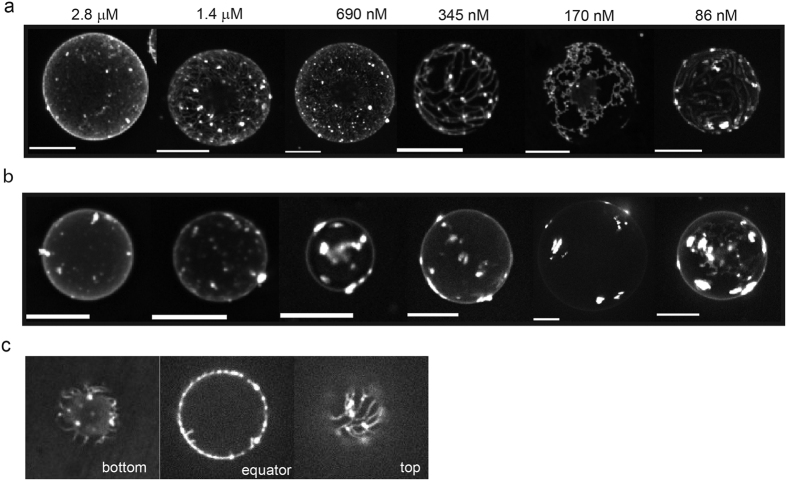
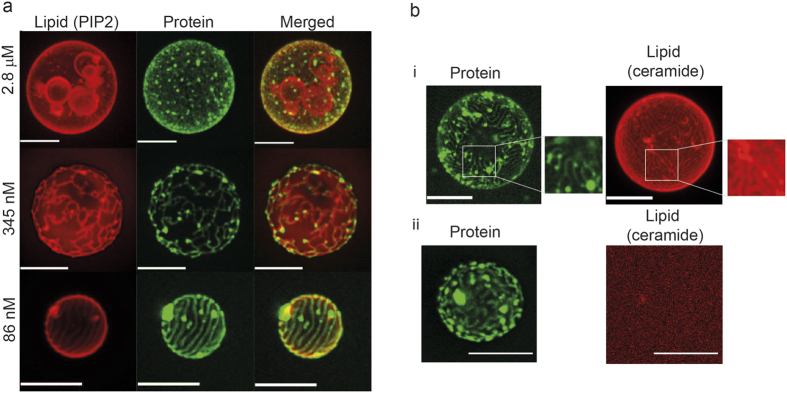
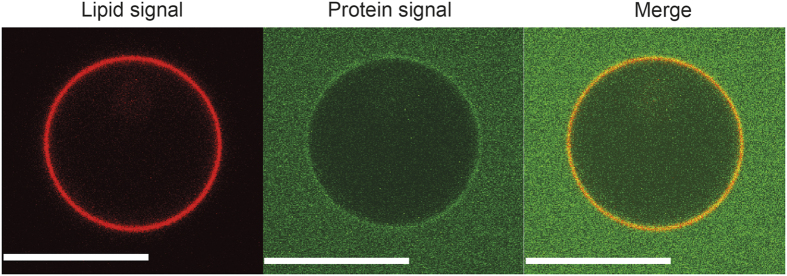
Matrix proteins from enveloped viruses play an important role in budding and stabilizing virus particles. In order to assess the role of the matrix protein M1 from influenza C virus (M1-C) in plasma membrane deformation, we have combined structural and in vitro reconstitution experiments with model membranes. We present the crystal structure of the N-terminal domain of M1-C and show by Small Angle X-Ray Scattering analysis that full-length M1-C folds into an elongated structure that associates laterally into ring-like or filamentous polymers. Using negatively charged giant unilamellar vesicles (GUVs), we demonstrate that M1-C full-length binds to and induces inward budding of membrane tubules with diameters that resemble the diameter of viruses. Membrane tubule formation requires the C-terminal domain of M1-C, corroborating its essential role for M1-C polymerization. Our results indicate that M1-C assembly on membranes constitutes the driving force for budding and suggest that M1-C plays a key role in facilitating viral egress.
Figures
References
-
- Homma M., Ohyama S. & Katagiri S. Age distribution of the antibody to type C influenza virus. Microbiol Immunol 26, 639–642 (1982). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
