

Three-Phase Coexistence in Lipid Membranes
- PMID: 28122217
- PMCID: PMC5266263
- DOI: 10.1016/j.bpj.2016.12.025
Three-Phase Coexistence in Lipid Membranes
Abstract
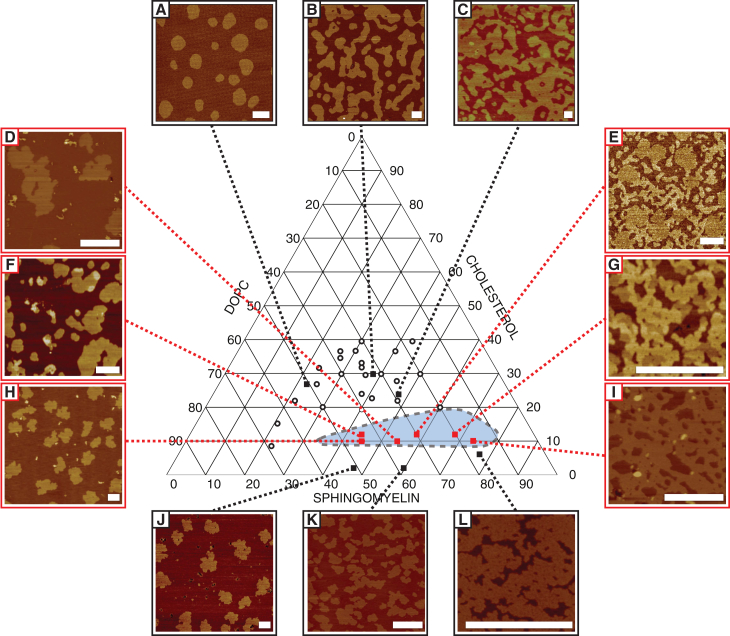
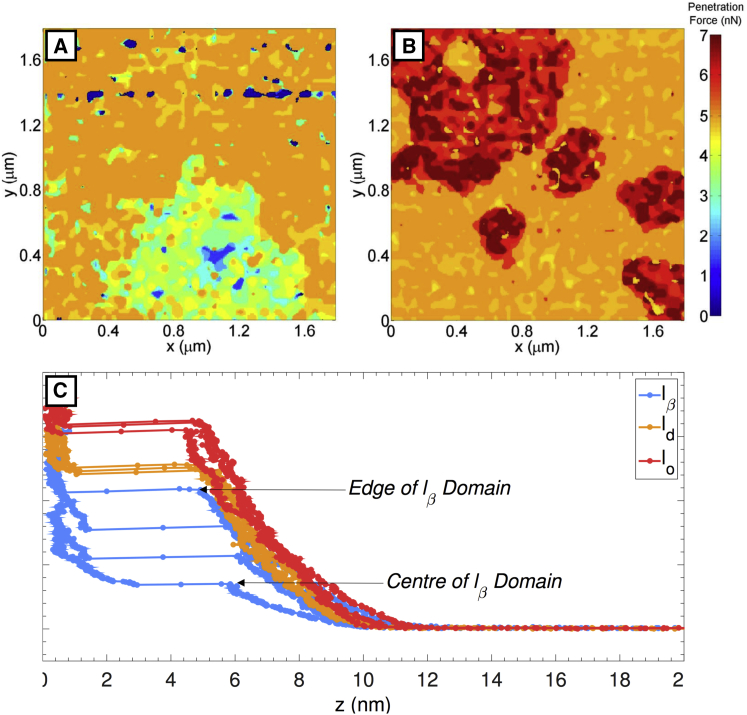
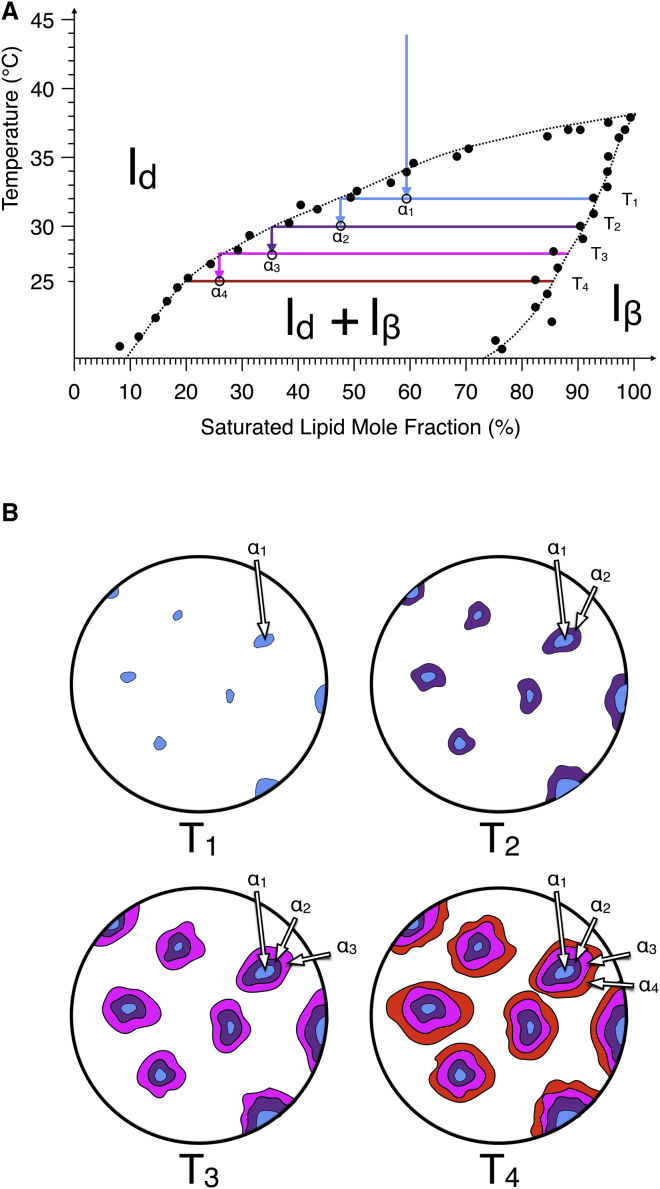
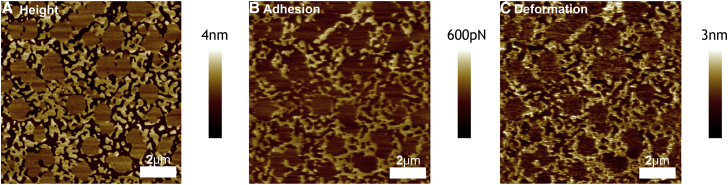
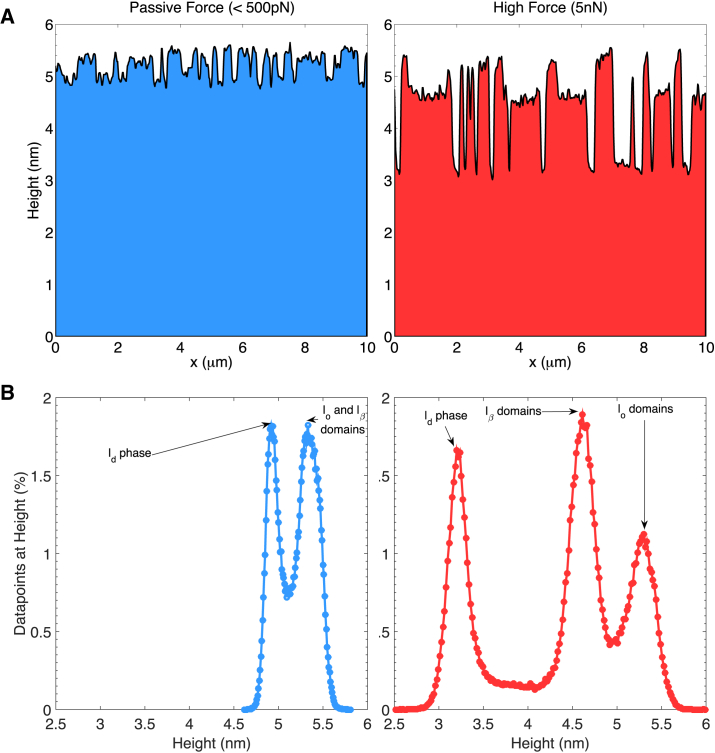
Phospholipid ternary systems are useful model systems for understanding lipid-lipid interactions and their influence on biological properties such as cell signaling and protein translocation. Despite extensive studies, there are still open questions relating to membrane phase behavior, particularly relating to a proposed state of three-phase coexistence, due to the difficulty in clearly distinguishing the three phases. We look in and around the region of the phase diagram where three phases are expected and use a combination of different atomic force microscopy (AFM) modes to present the first images of three coexisting lipid phases in biomimetic cell lipid membranes. Domains form through either nucleation or spinodal decomposition dependent upon composition, with some exhibiting both mechanisms in different domains simultaneously. Slow cooling rates are necessary to sufficiently separate mixtures with high proportions of lo and lβ phase. We probe domain heights and mechanical properties and demonstrate that the gel (lβ) domains have unusually low structural integrity in the three-phase region. This finding supports the hypothesis of a "disordered gel" state that has been proposed from NMR studies of similar systems, where the addition of small amounts of cholesterol was shown to disrupt the regular packing of the lβ state. We use NMR data from the literature on chain disorder in different mixtures and estimate an expected step height that is in excellent agreement with the AFM data. Alternatively, the disordered solid phase observed here and in the wider literature could be explained by the lβ phase being out of equilibrium, in a surface kinetically trapped state. This view is supported by the observation of unusual growth of nucleated domains, which we term "tree-ring growth," reflecting compositional heterogeneity in large disordered lβ phase domains.
Copyright © 2017 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures


References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
