

Synthetic RORγ agonists regulate multiple pathways to enhance antitumor immunity
- PMID: 28123897
- PMCID: PMC5215247
- DOI: 10.1080/2162402X.2016.1254854
Synthetic RORγ agonists regulate multiple pathways to enhance antitumor immunity
Abstract
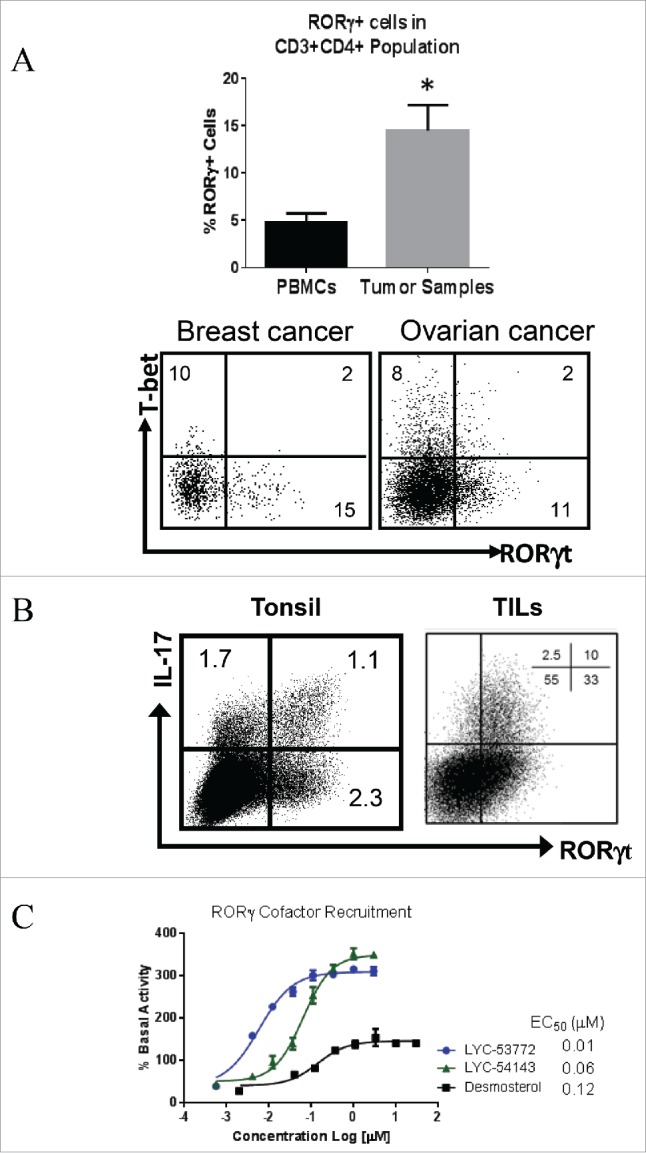
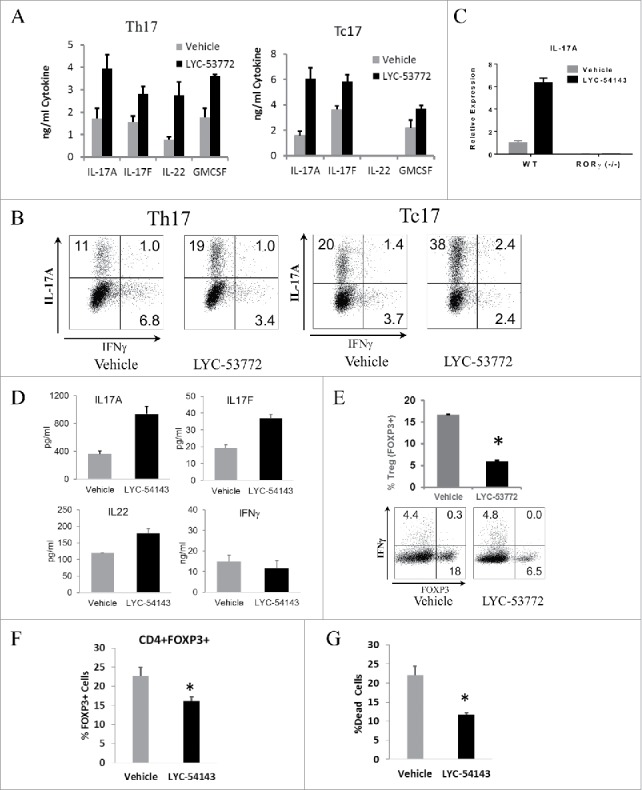
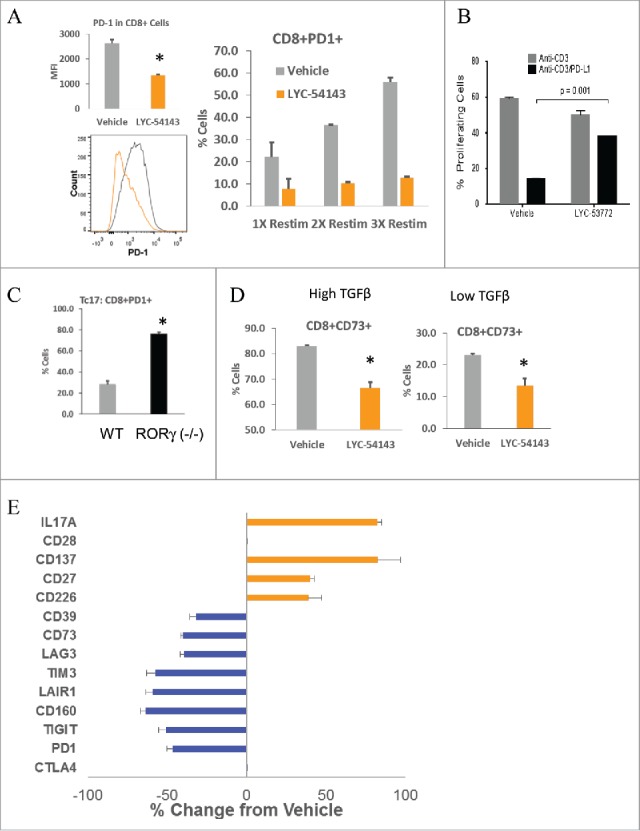
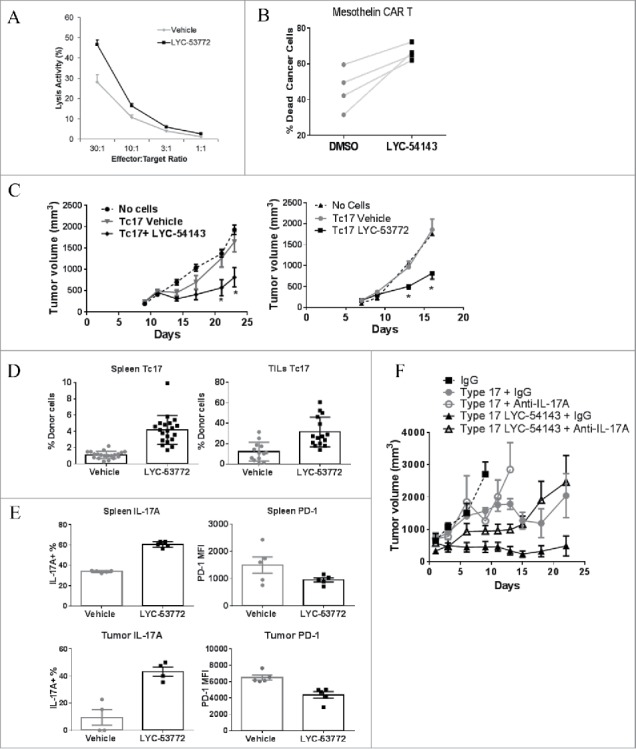
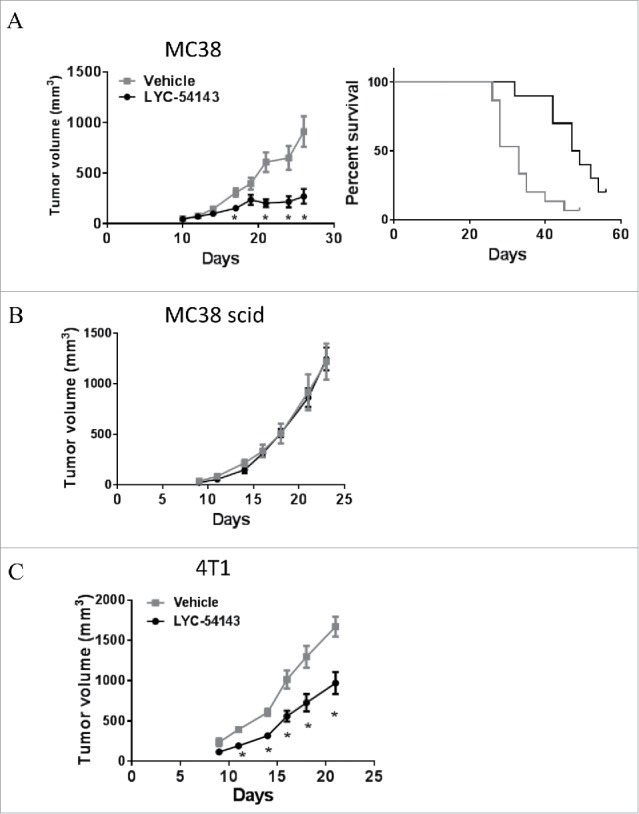
RORγt is the key transcription factor controlling the development and function of CD4+ Th17 and CD8+ Tc17 cells. Across a range of human tumors, about 15% of the CD4+ T cell fraction in tumor-infiltrating lymphocytes are RORγ+ cells. To evaluate the role of RORγ in antitumor immunity, we have identified synthetic, small molecule agonists that selectively activate RORγ to a greater extent than the endogenous agonist desmosterol. These RORγ agonists enhance effector function of Type 17 cells by increasing the production of cytokines/chemokines such as IL-17A and GM-CSF, augmenting expression of co-stimulatory receptors like CD137, CD226, and improving survival and cytotoxic activity. RORγ agonists also attenuate immunosuppressive mechanisms by curtailing Treg formation, diminishing CD39 and CD73 expression, and decreasing levels of co-inhibitory receptors including PD-1 and TIGIT on tumor-reactive lymphocytes. The effects of RORγ agonists were not observed in RORγ-/- T cells, underscoring the selective on-target activity of the compounds. In vitro treatment of tumor-specific T cells with RORγ agonists, followed by adoptive transfer to tumor-bearing mice is highly effective at controlling tumor growth while improving T cell survival and maintaining enhanced IL-17A and reduced PD-1 in vivo. The in vitro effects of RORγ agonists translate into single agent, immune system-dependent, antitumor efficacy when compounds are administered orally in syngeneic tumor models. RORγ agonists integrate multiple antitumor mechanisms into a single therapeutic that both increases immune activation and decreases immune suppression resulting in robust inhibition of tumor growth. Thus, RORγ agonists represent a novel immunotherapy approach for cancer.
Keywords: Adoptive cell therapy; PD-1; RORγ; Tc17; Th17; co-inhibitory receptors; co-stimulatory receptors; immunotherapy.
Figures
References
-
- Cua DJ, Tato CM. Innate IL-17-producing cells: the sentinels of the immune system. Nat Rev Immunol 2010; 10:479-89; PMID:20559326; http://dx.doi.org/ 10.1038/nri2800 - DOI - PubMed
-
- Ivanov II, McKenzie BS, Zhou L, Tadokoro CE, Lepelley A, Lafaille JJ, Cua DJ, Littman DR. The orphan nuclear receptor RORgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell 2006; 126:1121-33; PMID:16990136; http://dx.doi.org/ 10.1016/j.cell.2006.07.035 - DOI - PubMed
-
- Codarri L, Gyulveszi G, Tosevski V, Hesske L, Fontana A, Magnenat L, Suter T, Becher B. RORgammat drives production of the cytokine GM-CSF in helper T cells, which is essential for the effector phase of autoimmune neuroinflammation. Nat Immunol 2011; 12:560-7; PMID:21516112; http://dx.doi.org/ 10.1038/ni.2027 - DOI - PubMed
-
- Manel N, Unutmaz D, Littman DR. The differentiation of human T(H)-17 cells requires transforming growth factor-β and induction of the nuclear receptor RORgammat. Nat Immunol 2008; 9:641-9; PMID:18454151; http://dx.doi.org/ 10.1038/ni.1610 - DOI - PMC - PubMed
-
- Dong C. Differentiation and function of pro-inflammatory Th17 cells. Microbes Infect / Institut Pasteur 2009; 11:584-8; PMID:19371793; http://dx.doi.org/10875923 10.1016/j.micinf.2009.04.001 - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials