

Toxoplasma gondii GRA7-Targeted ASC and PLD1 Promote Antibacterial Host Defense via PKCα
- PMID: 28125719
- PMCID: PMC5268361
- DOI: 10.1371/journal.ppat.1006126
Toxoplasma gondii GRA7-Targeted ASC and PLD1 Promote Antibacterial Host Defense via PKCα
Abstract
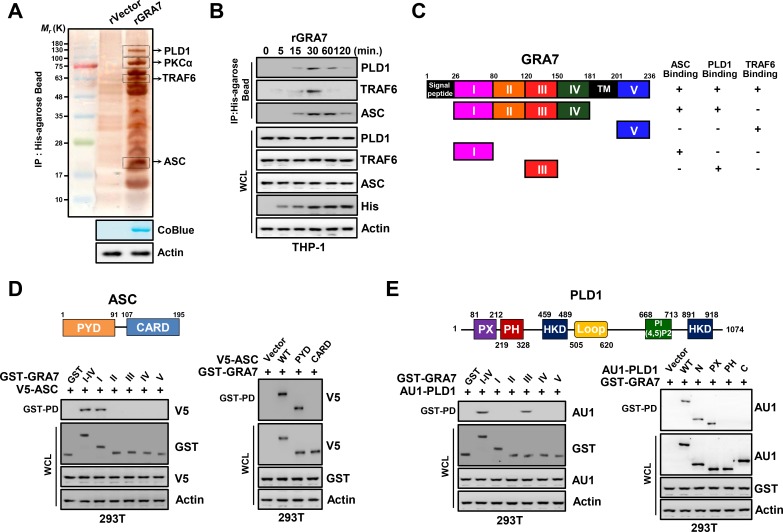
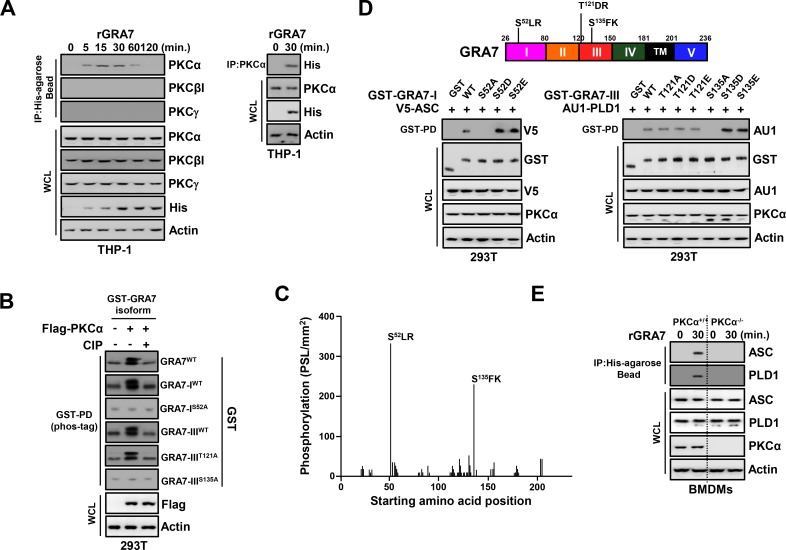
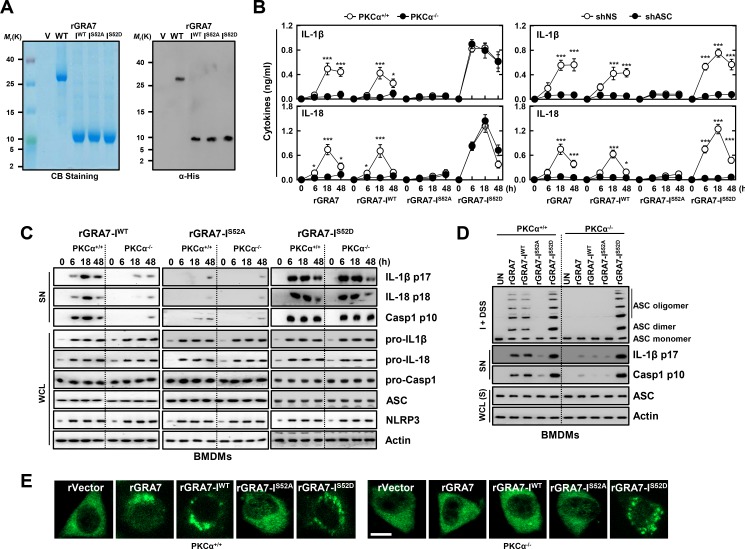
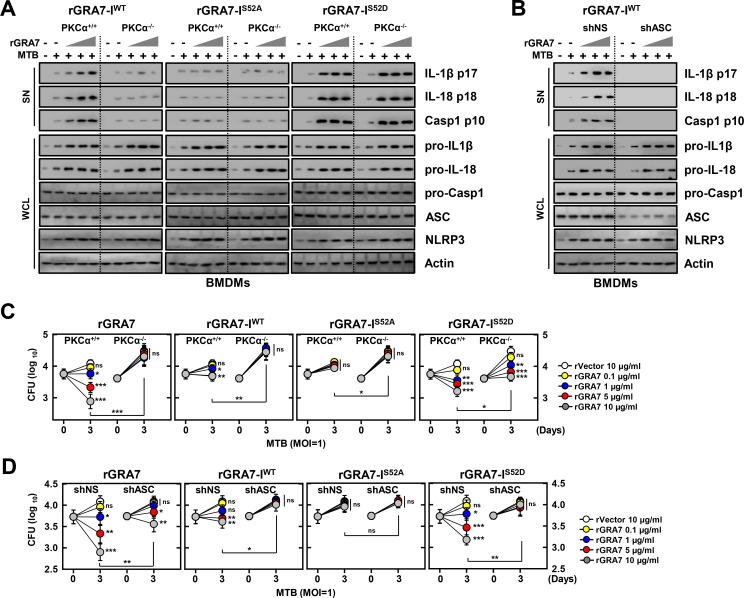
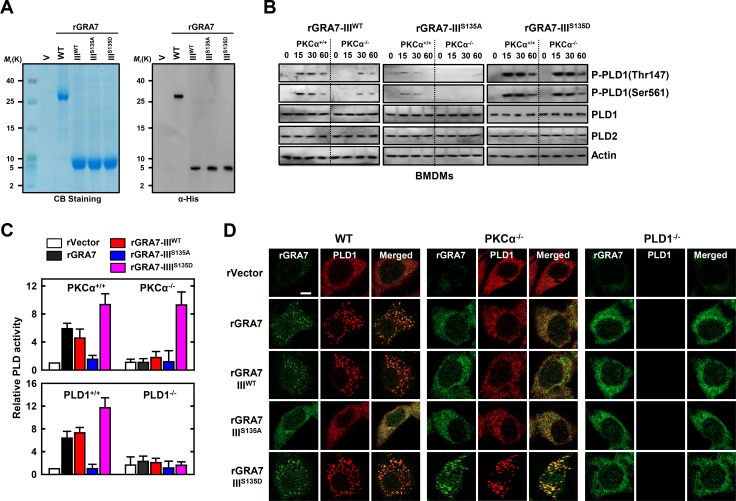
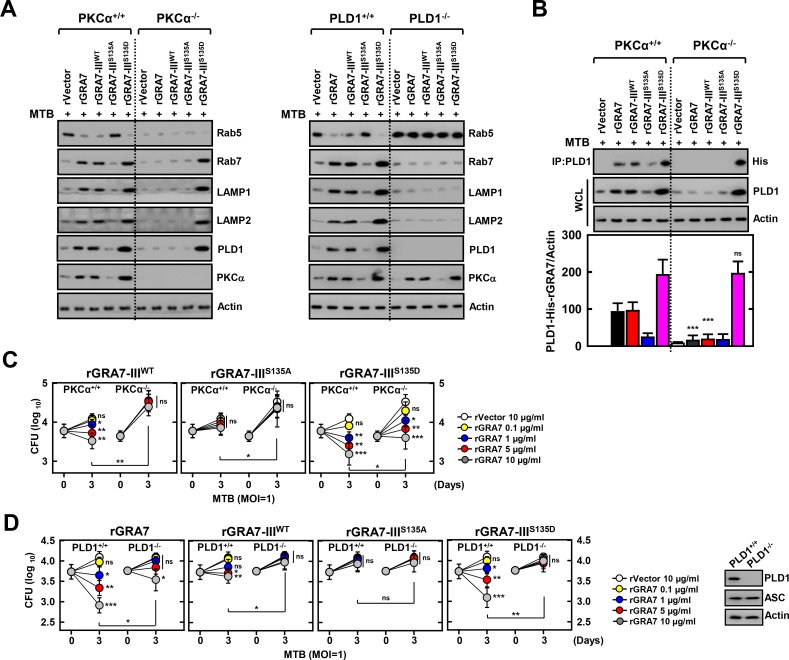
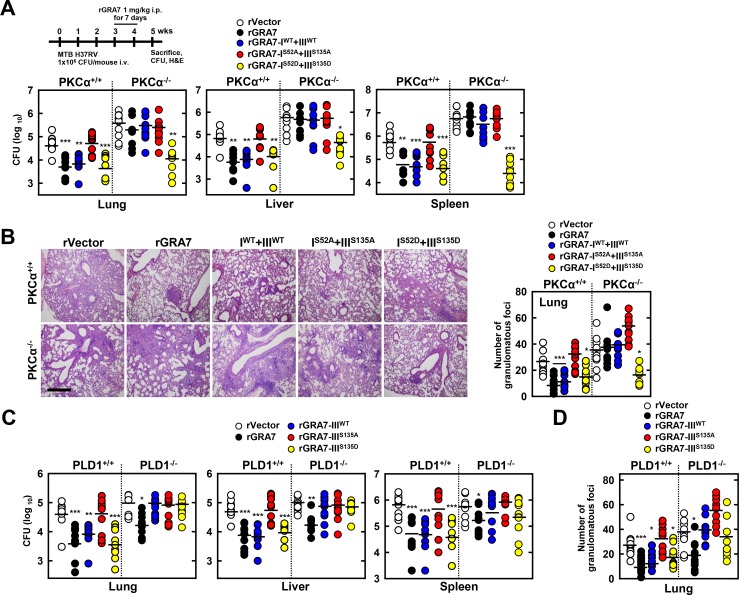
Tuberculosis is a global health problem and at least one-third of the world's population is infected with Mycobacterium tuberculosis (MTB). MTB is a successful pathogen that enhances its own intracellular survival by inhibiting inflammation and arresting phago-lysosomal fusion. We previously demonstrated that Toxoplasma gondii (T. gondii) dense granule antigen (GRA) 7 interacts with TNF receptor-associated factor 6 via Myeloid differentiation primary response gene 88, enabling innate immune responses in macrophages. To extend these studies, we found that GRA7 interacts with host proteins involved in antimicrobial host defense mechanisms as a therapeutic strategy for tuberculosis. Here, we show that protein kinase C (PKC)α-mediated phosphorylation of T. gondii GRA7-I (Ser52) regulates the interaction of GRA7 with PYD domain of apoptosis-associated speck-like protein containing a carboxy-terminal CARD, which is capable of oligomerization and inflammasome activation can lead to antimicrobial defense against MTB. Furthermore, GRA7-III interacted with the PX domain of phospholipase D1, facilitating its enzyme activity, phago-lysosomal maturation, and subsequent antimicrobial activity in a GRA7-III (Ser135) phosphorylation-dependent manner via PKCα. Taken together, these results underscore a previously unrecognized role of GRA7 in modulating antimicrobial host defense mechanism during mycobacterial infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Comment in
-
Advancing host-directed therapy for tuberculosis.Microb Cell. 2017 Mar 2;4(3):105-107. doi: 10.15698/mic2017.03.565. Microb Cell. 2017. PMID: 28357397 Free PMC article.
References
-
- (2015) World Health Organization releases 2015 global report on tuberculosis. Breathe 11: 244–244.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
