

Lipids regulate the hydrolysis of membrane bound glucosylceramide by lysosomal β-glucocerebrosidase
- PMID: 28126847
- PMCID: PMC5335586
- DOI: 10.1194/jlr.M073510
Lipids regulate the hydrolysis of membrane bound glucosylceramide by lysosomal β-glucocerebrosidase
Abstract
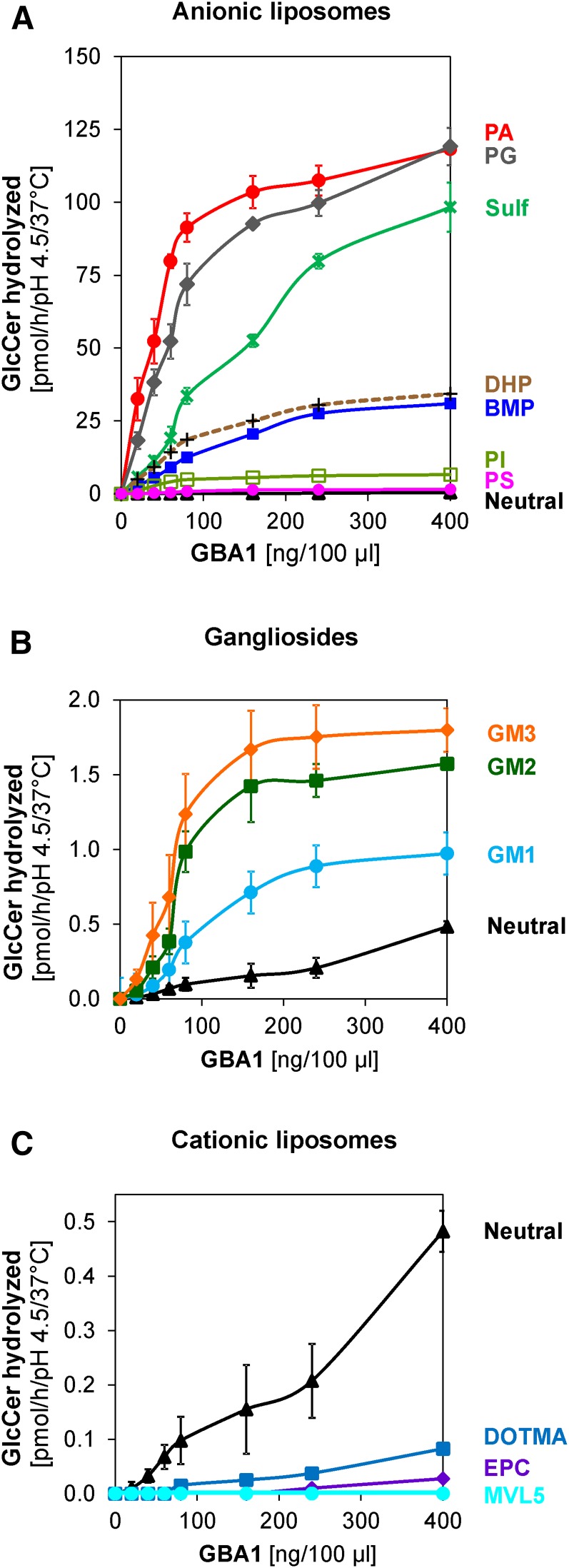
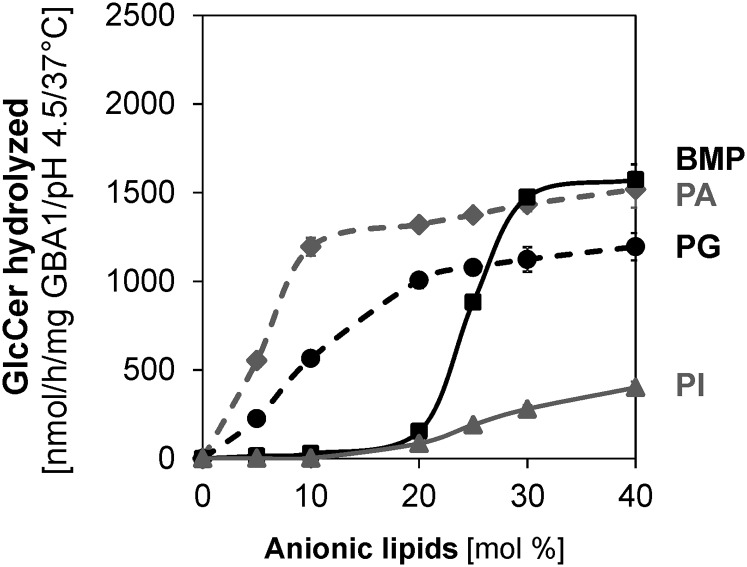
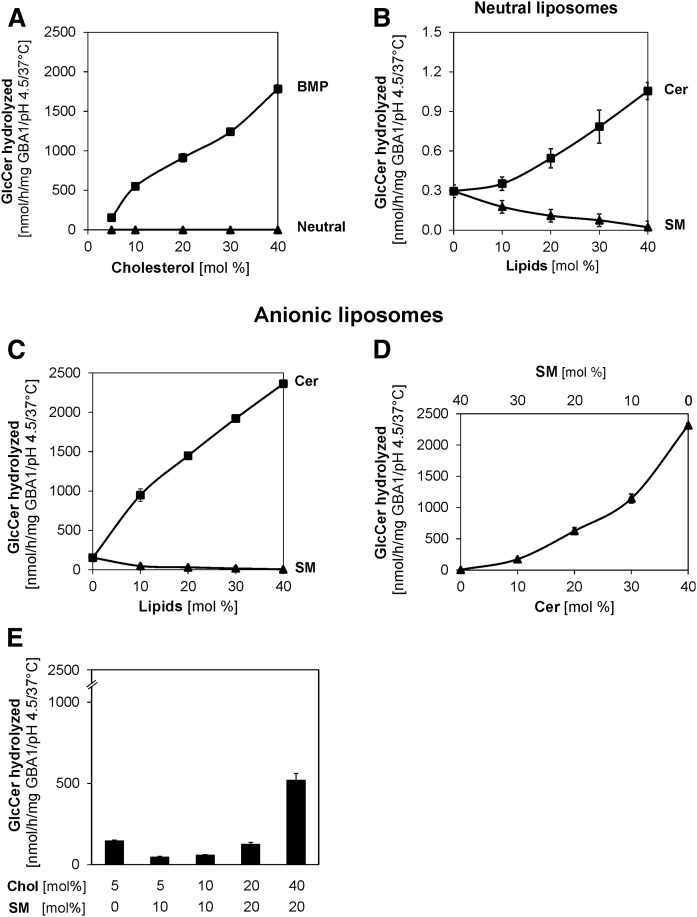
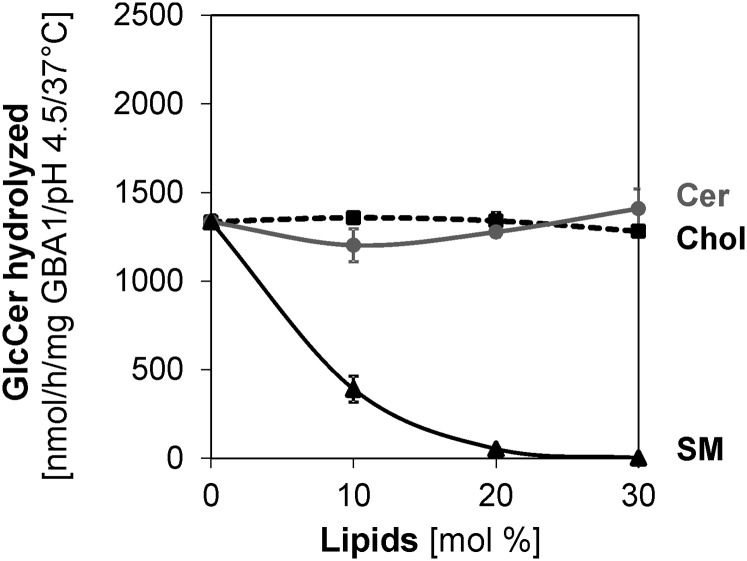
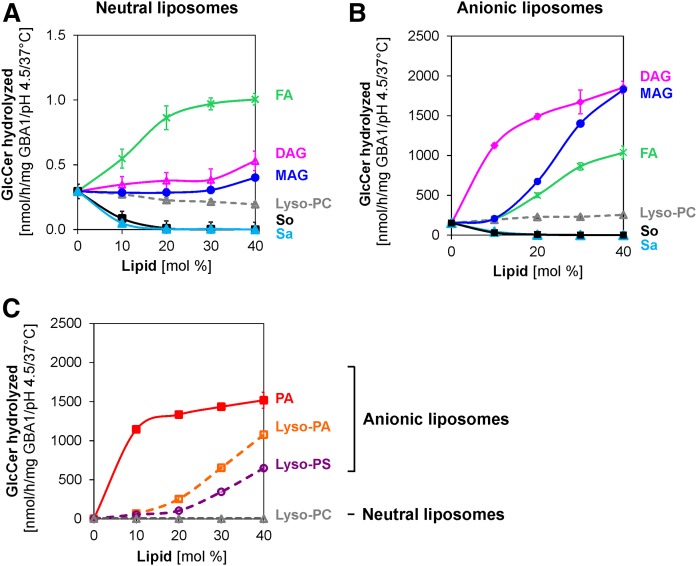
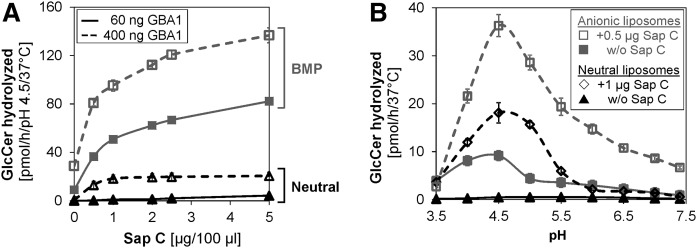
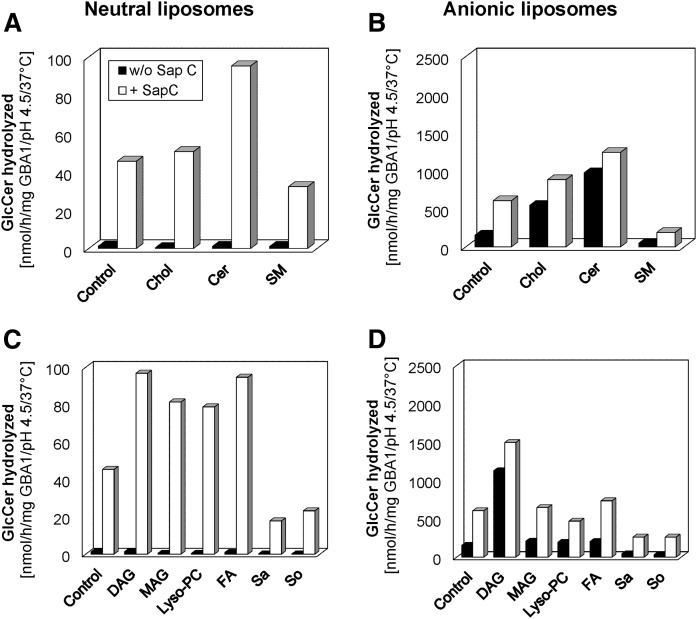
Glucosylceramide (GlcCer) is the primary storage lipid in the lysosomes of Gaucher patients and a secondary one in Niemann-Pick disease types A, B, and C. The regulatory roles of lipids on the hydrolysis of membrane bound GlcCer by lysosomal β-glucocerebrosidase (GBA1) was probed using a detergent-free liposomal assay. The degradation rarely occurs at uncharged liposomal surfaces in the absence of saposin (Sap) C. However, anionic lipids stimulate GlcCer hydrolysis at low pH by up to 1,000-fold depending on the nature and position of the negative charges in their head groups while cationic lipids inhibit the degradation, thus showing the importance of electrostatic interactions between the polycationic GBA1 and the negatively charged vesicle surfaces at low pH. Ceramide, fatty acids, monoacylglycerol, and diacylglycerol also stimulate GlcCer hydrolysis while SM, sphingosine, and sphinganine play strong inhibitory roles, thereby explaining the secondary storage of GlcCer in Niemann-Pick diseases. Surprisingly, cholesterol stimulates GlcCer degradation in the presence of bis(monoacylglycero)phosphate (BMP). Sap C strongly stimulates GlcCer hydrolysis even in the absence of BMP and the regulatory roles of the intraendolysosomal lipids on its activity is discussed. Our data suggest that these strong modifiers of GlcCer hydrolysis affect the genotype-phenotype correlation in several cases of Gaucher patients independent of the types.
Keywords: Gaucher disease; acylglycerols; anionic phospholipids; cationic lipids; cholesterol; electrostatic interaction; fatty acids; lysophospholipids; sphingomyelin; sphingosine.
Copyright © 2017 by the American Society for Biochemistry and Molecular Biology, Inc.
Figures

References
-
- Schulze H., Kolter T., and Sandhoff K.. 2009. Principles of lysosomal membrane degradation: Cellular topology and biochemistry of lysosomal lipid degradation. Biochim. Biophys. Acta. 1793: 674–683. - PubMed
-
- Brady R. O., Kanfer J. N., and Shapiro D.. 1965. Metabolism of glucocerebrosides. II. Evidence of an enzymatic deficiency in Gaucher’s disease. Biochem. Biophys. Res. Commun. 18: 221–225. - PubMed
-
- Sasaki T. 1990. Glycolipid transfer protein and intracellular traffic of glucosylceramide. Experientia. 46: 611–616. - PubMed
-
- Möbius W., Herzog V., Sandhoff K., and Schwarzmann G.. 1999. Intracellular distribution of a biotin-labeled ganglioside, GM1, by immunoelectron microscopy after endocytosis in fibroblasts. J. Histochem. Cytochem. 47: 1005–1014. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
