

DNA Double-Strand Break Resection Occurs during Non-homologous End Joining in G1 but Is Distinct from Resection during Homologous Recombination
- PMID: 28132842
- PMCID: PMC5316416
- DOI: 10.1016/j.molcel.2016.12.016
DNA Double-Strand Break Resection Occurs during Non-homologous End Joining in G1 but Is Distinct from Resection during Homologous Recombination
Abstract
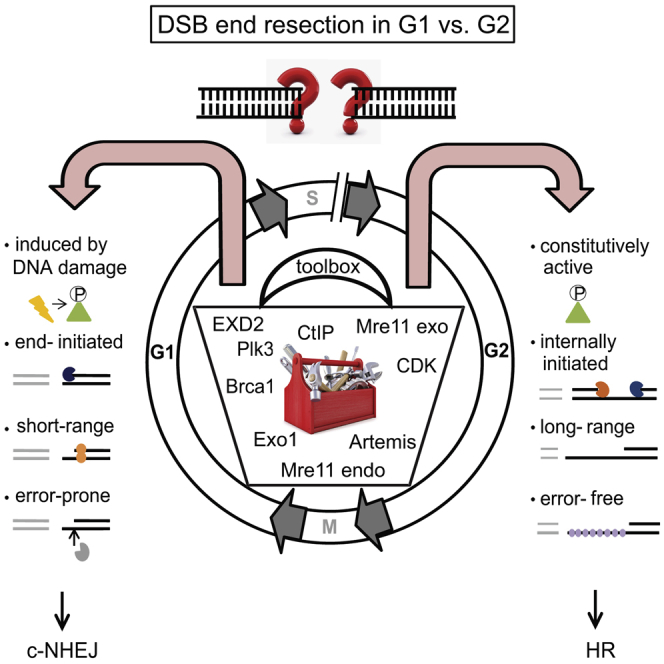
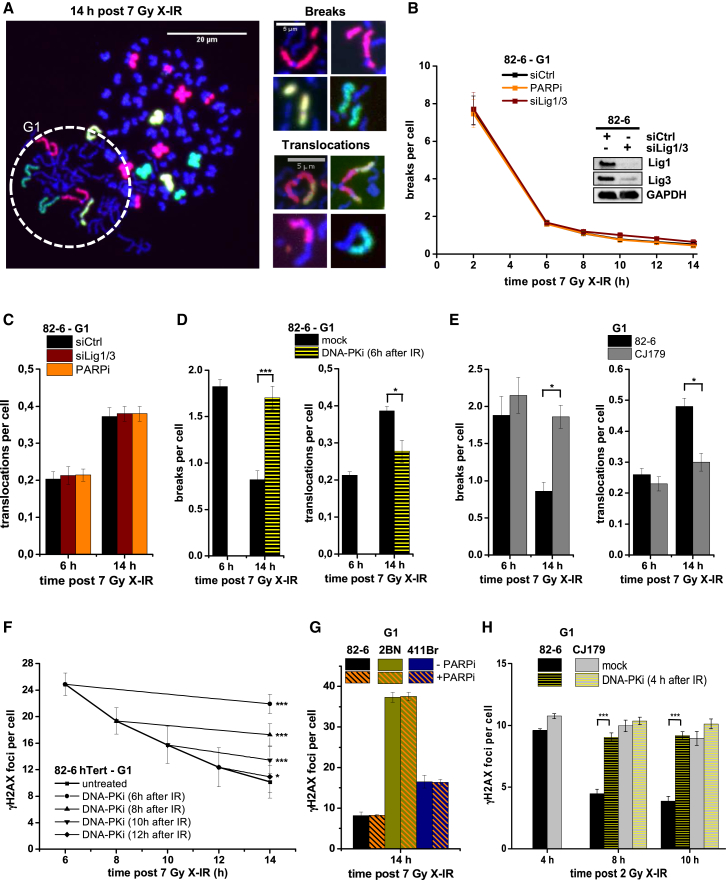
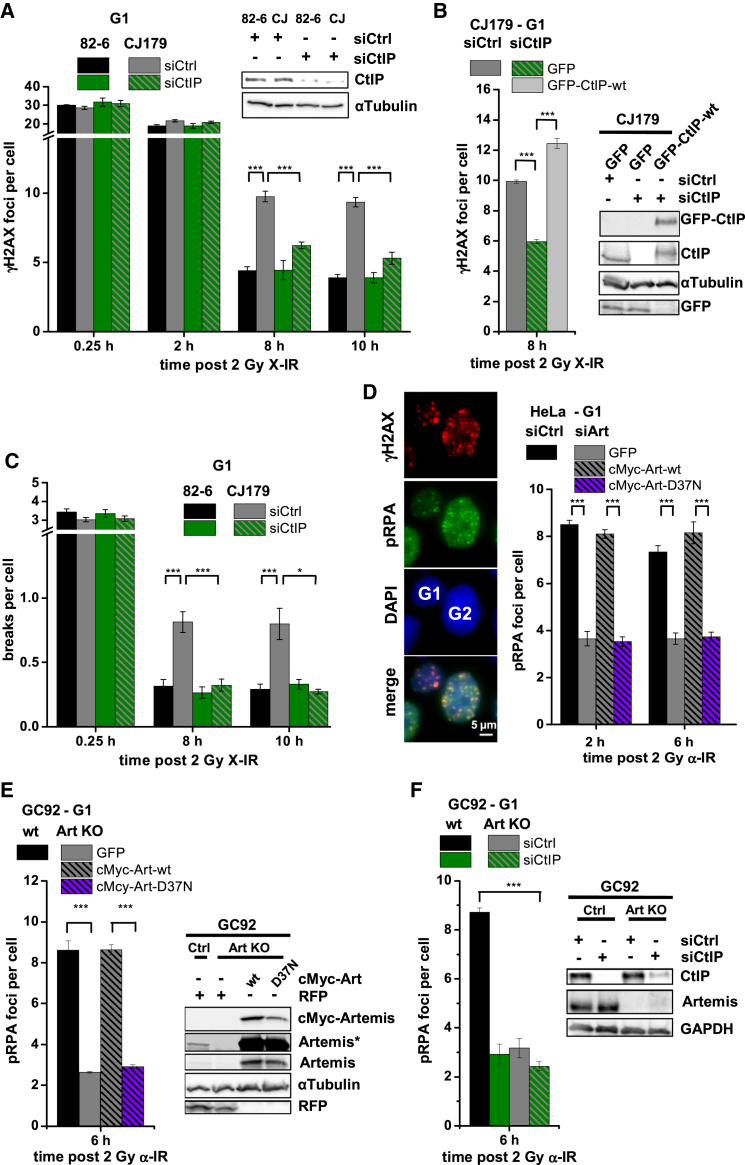
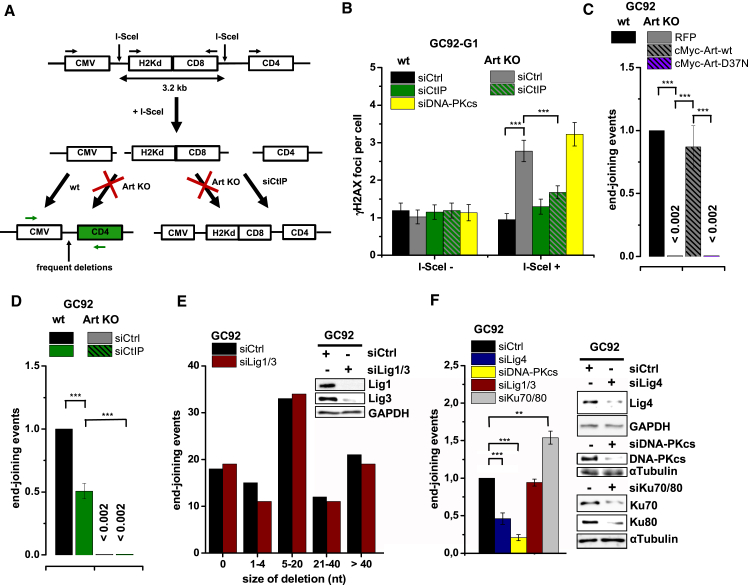
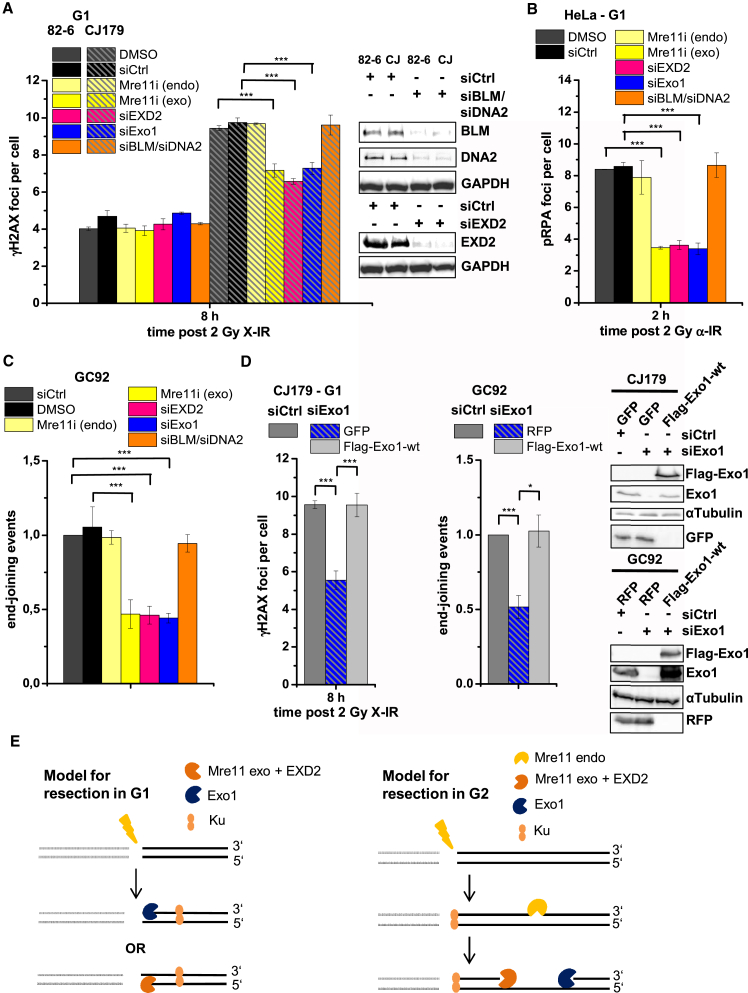
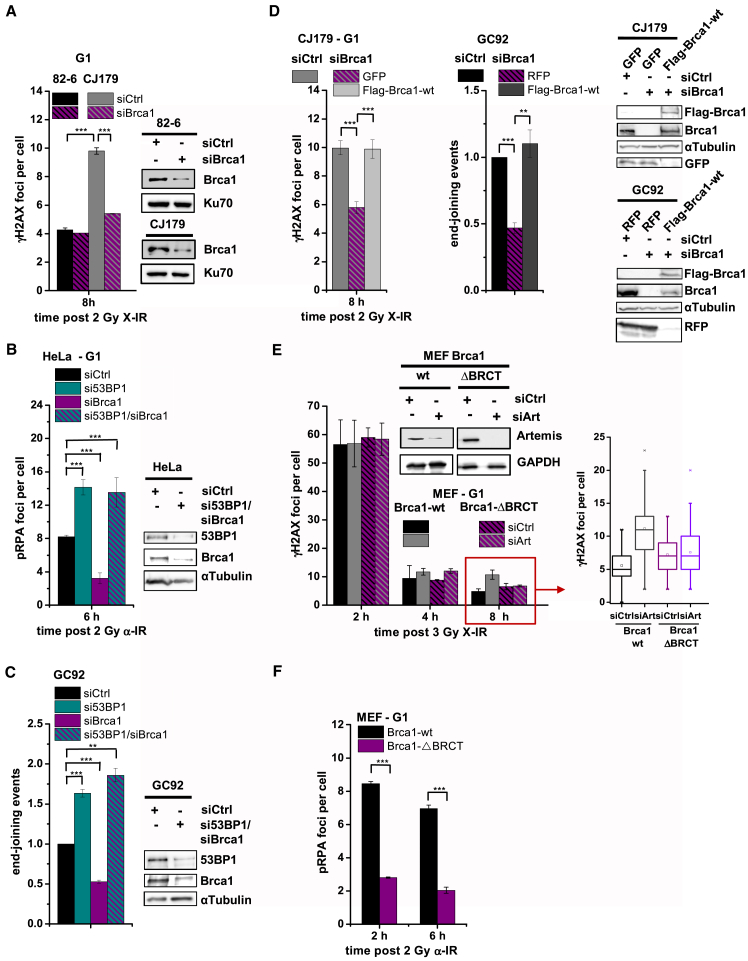
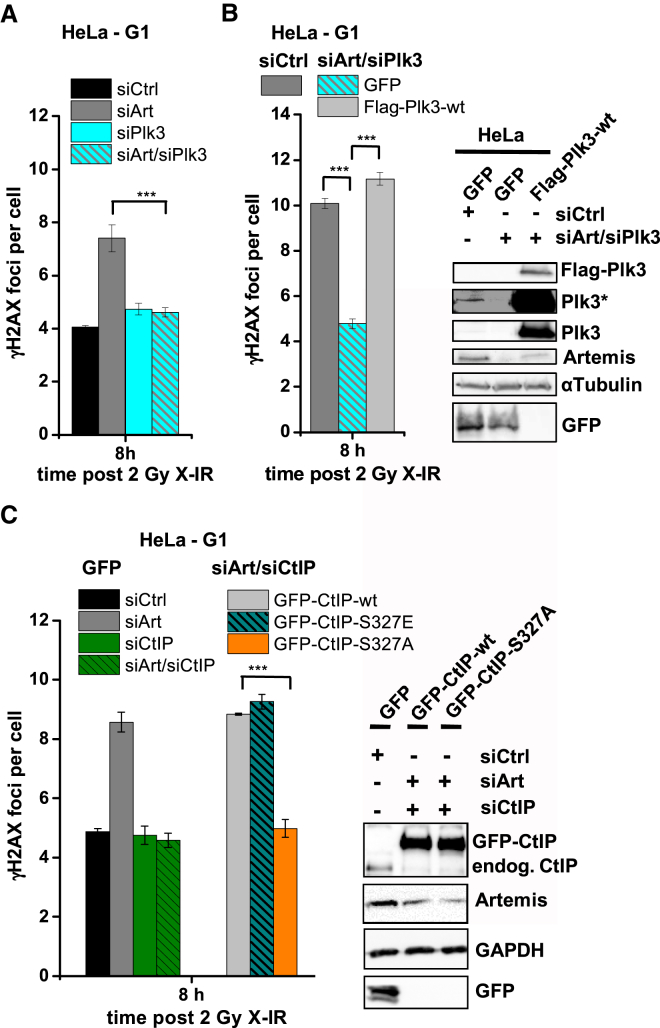
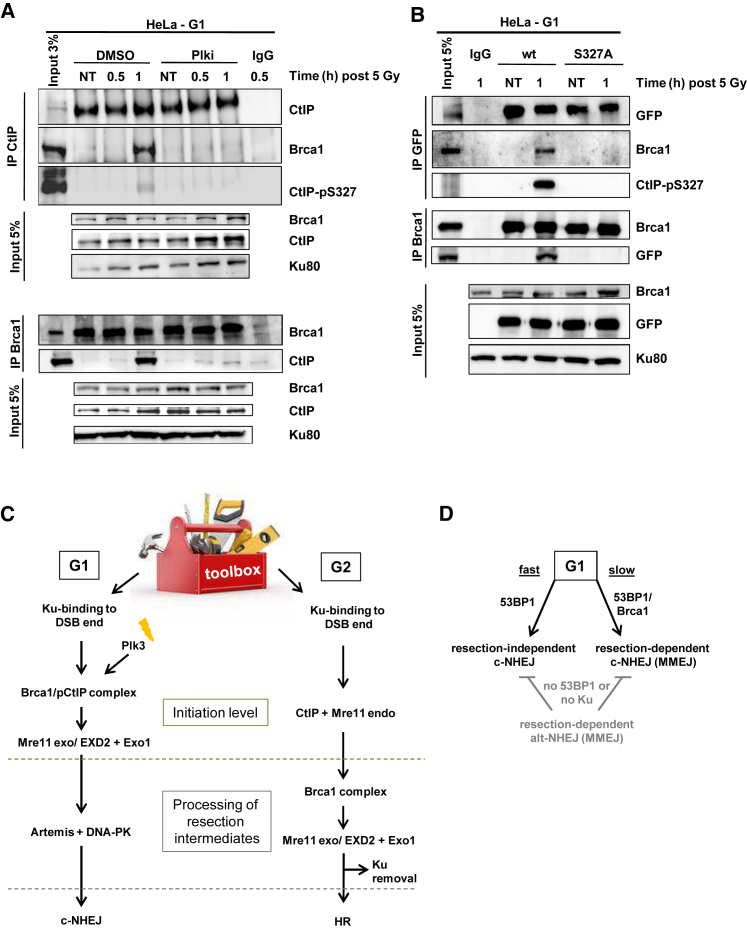
Canonical non-homologous end joining (c-NHEJ) repairs DNA double-strand breaks (DSBs) in G1 cells with biphasic kinetics. We show that DSBs repaired with slow kinetics, including those localizing to heterochromatic regions or harboring additional lesions at the DSB site, undergo resection prior to repair by c-NHEJ and not alt-NHEJ. Resection-dependent c-NHEJ represents an inducible process during which Plk3 phosphorylates CtIP, mediating its interaction with Brca1 and promoting the initiation of resection. Mre11 exonuclease, EXD2, and Exo1 execute resection, and Artemis endonuclease functions to complete the process. If resection does not commence, then repair can ensue by c-NHEJ, but when executed, Artemis is essential to complete resection-dependent c-NHEJ. Additionally, Mre11 endonuclease activity is dispensable for resection in G1. Thus, resection in G1 differs from the process in G2 that leads to homologous recombination. Resection-dependent c-NHEJ significantly contributes to the formation of deletions and translocations in G1, which represent important initiating events in carcinogenesis.
Keywords: DNA double-strand breaks; non-homologous end joining; nucleases; resection.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
