

ON or OFF?: Modulating the N-Methyl-D-Aspartate Receptor in Major Depression
- PMID: 28133445
- PMCID: PMC5233677
- DOI: 10.3389/fnmol.2016.00169
ON or OFF?: Modulating the N-Methyl-D-Aspartate Receptor in Major Depression
Abstract
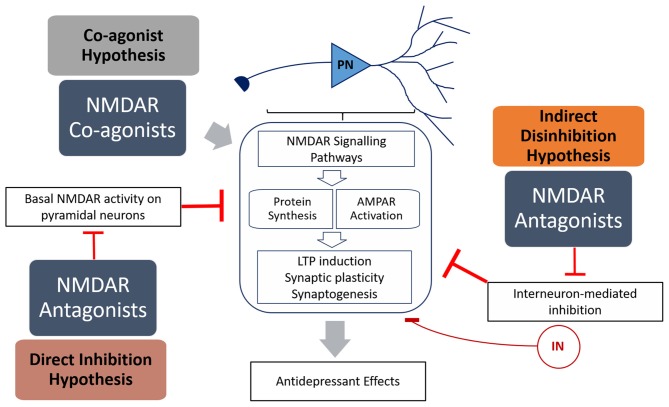
Since the discovery that a single dose of ketamine, an N-methyl-D-aspartate receptor (NMDAR) antagonist, had rapid and long-lasting antidepressant effects, there has been increased interest in using NMDAR modulators in the pharmacotherapy of depression. Ketamine's efficacy seems to imply that depression is a disorder of NMDAR hyperfunctionality. However, studies showing that not all NMDAR antagonists are able to act as antidepressants challenge this notion. Furthermore, NMDAR co-agonists have also been gaining attention as possible treatments. Co-agonists such as D-serine and sarcosine have shown efficacy in both pre-clinical models and human trials. This raises the question of how both NMDAR antagonists and agonists are able to have converging behavioral effects. Here we critically review the evidence and proposed therapeutic mechanisms for both NMDAR antagonists and agonists, and collate several theories on how both activation and inhibition of NMDARs appear to have antidepressant effects.
Keywords: NMDAR antagonist; depression; glycine site; mTOR; subunit.
Figures

References
-
- Andreasen J. T., Gynther M., Rygaard A., Bogelund T., Nielsen S. D., Clausen R. P., et al. . (2013). Does increasing the ratio of AMPA-to-NMDA receptor mediated neurotransmission engender antidepressant action? Studies in the mouse forced swim and tail suspension tests. Neurosci. Lett. 546, 6–10. 10.1016/j.neulet.2013.04.045 - DOI - PubMed
-
- van Berckel B. N., Evenblij C. N., van Loon B. J., Maas M. F., van der Geld M. A., Wynne H. J., et al. . (1999). D-cycloserine increases positive symptoms in chronic schizophrenic patients when administered in addition to antipsychotics: a double-blind, parallel, placebo-controlled study. Neuropsychopharmacology 21, 203–210. 10.1016/s0893-133x(99)00014-7 - DOI - PubMed
-
- Berger A. J., Dieudonné S., Ascher P. (1998). Glycine uptake governs glycine site occupancy at NMDA receptors of excitatory synapses. J. Neurophysiol. 80, 3336–3340. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous