

Portal protein functions akin to a DNA-sensor that couples genome-packaging to icosahedral capsid maturation
- PMID: 28134243
- PMCID: PMC5290284
- DOI: 10.1038/ncomms14310
Portal protein functions akin to a DNA-sensor that couples genome-packaging to icosahedral capsid maturation
Abstract
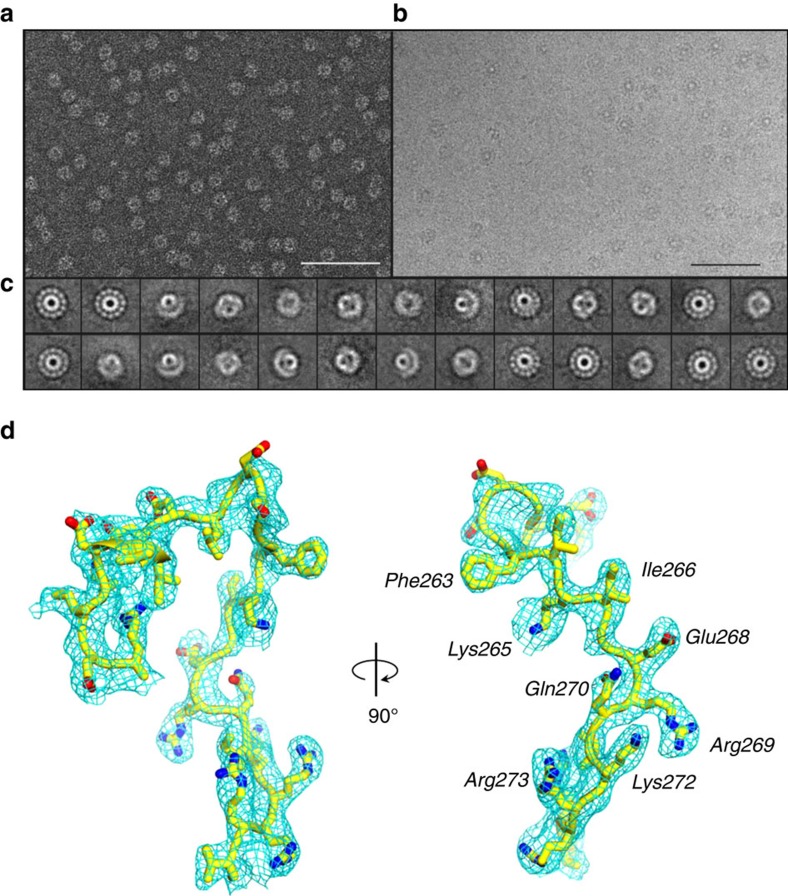
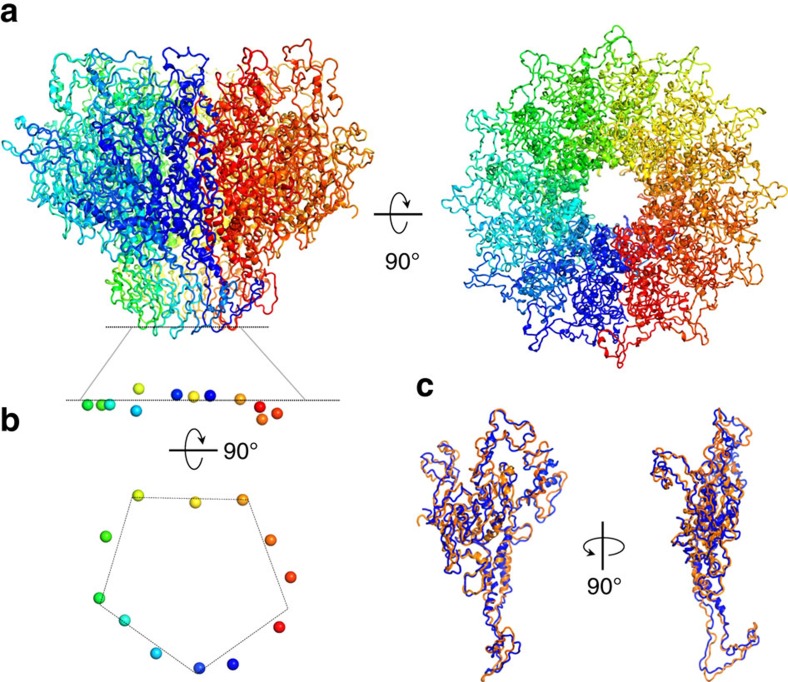
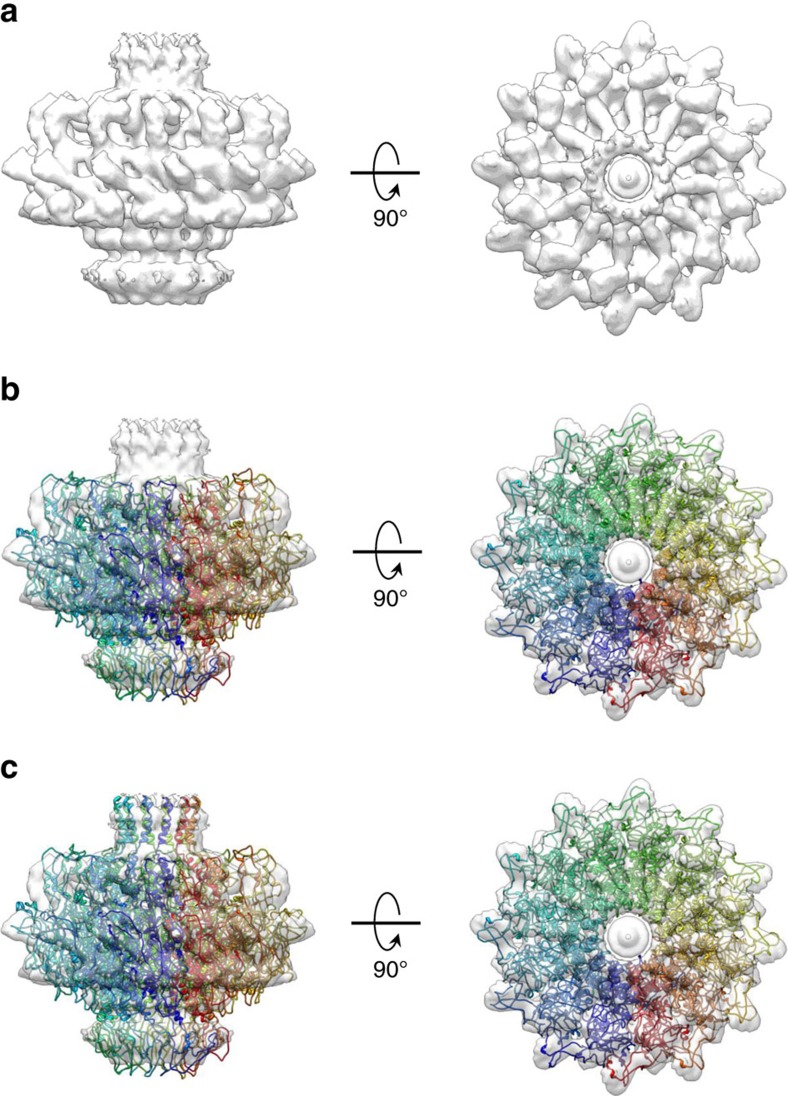
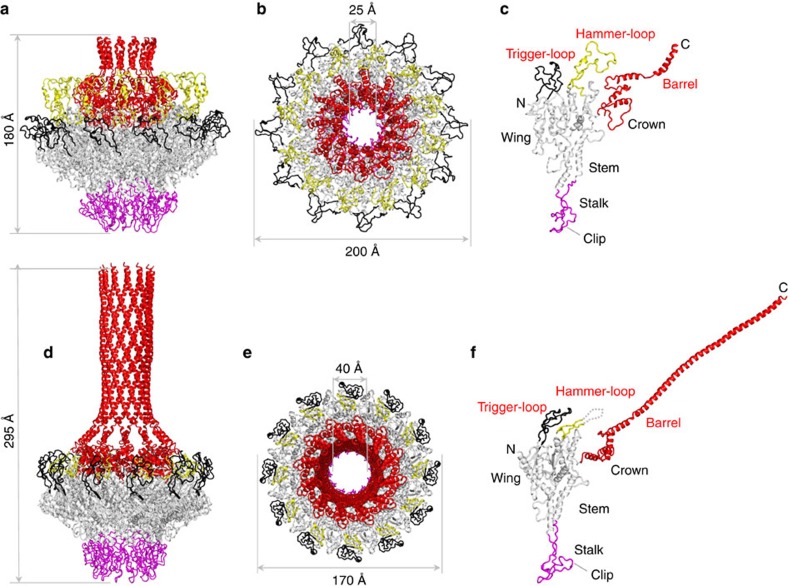
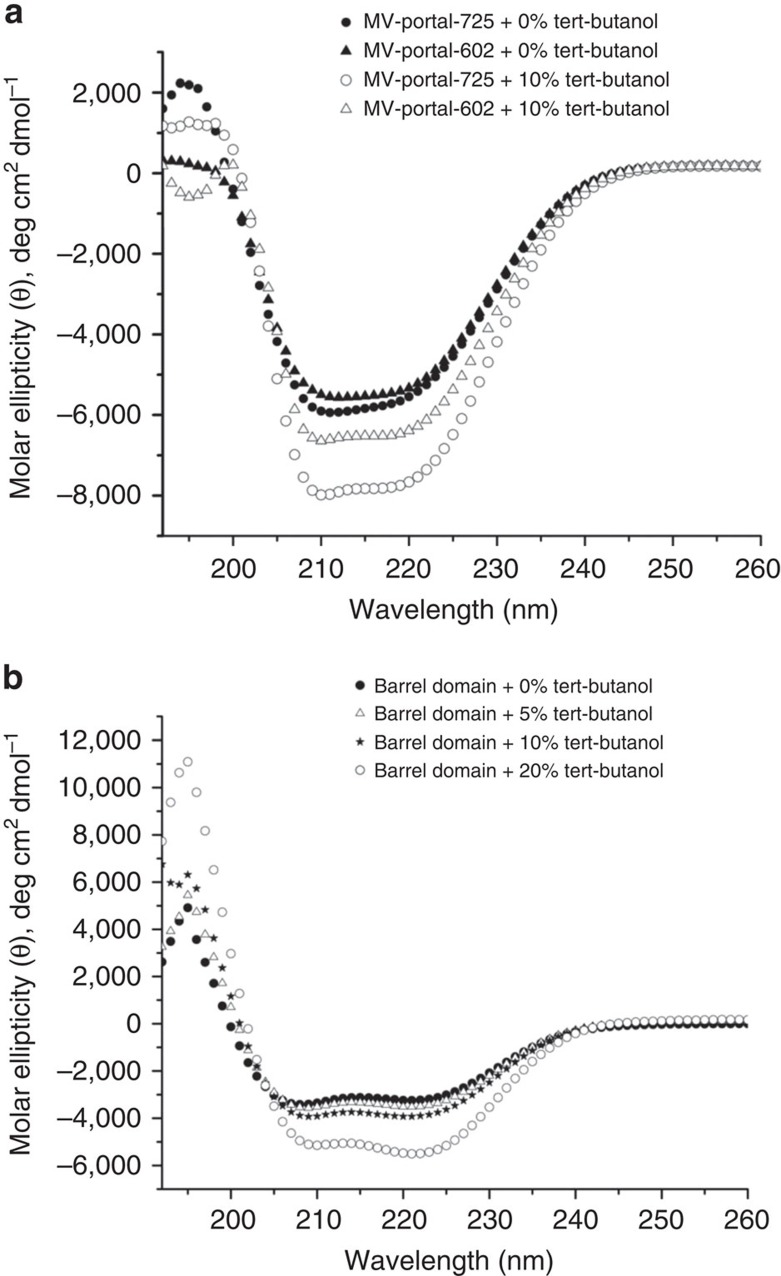
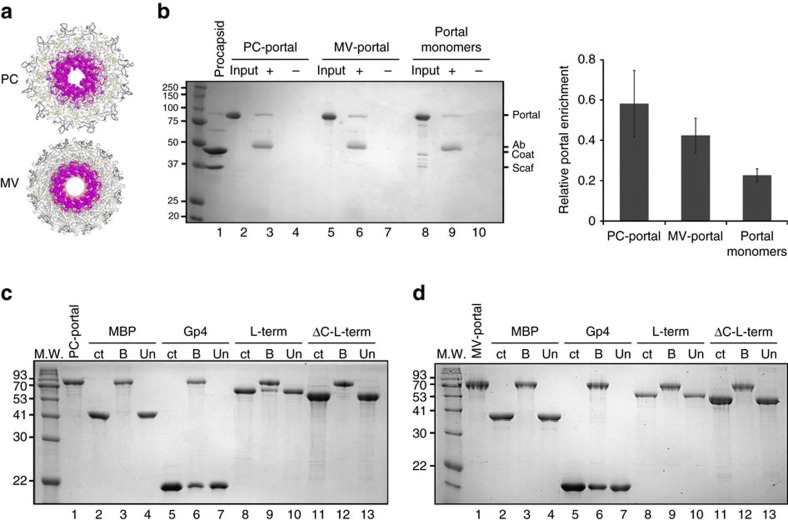
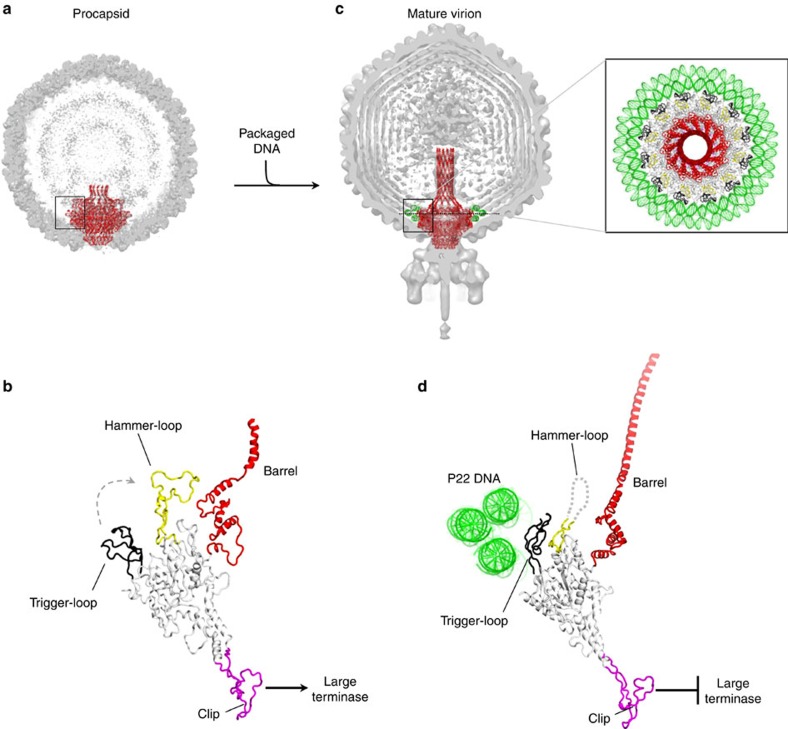
Tailed bacteriophages and herpesviruses assemble infectious particles via an empty precursor capsid (or 'procapsid') built by multiple copies of coat and scaffolding protein and by one dodecameric portal protein. Genome packaging triggers rearrangement of the coat protein and release of scaffolding protein, resulting in dramatic procapsid lattice expansion. Here, we provide structural evidence that the portal protein of the bacteriophage P22 exists in two distinct dodecameric conformations: an asymmetric assembly in the procapsid (PC-portal) that is competent for high affinity binding to the large terminase packaging protein, and a symmetric ring in the mature virion (MV-portal) that has negligible affinity for the packaging motor. Modelling studies indicate the structure of PC-portal is incompatible with DNA coaxially spooled around the portal vertex, suggesting that newly packaged DNA triggers the switch from PC- to MV-conformation. Thus, we propose the signal for termination of 'Headful Packaging' is a DNA-dependent symmetrization of portal protein.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
