

The paranodal cytoskeleton clusters Na+ channels at nodes of Ranvier
- PMID: 28134616
- PMCID: PMC5279941
- DOI: 10.7554/eLife.21392
The paranodal cytoskeleton clusters Na+ channels at nodes of Ranvier
Abstract
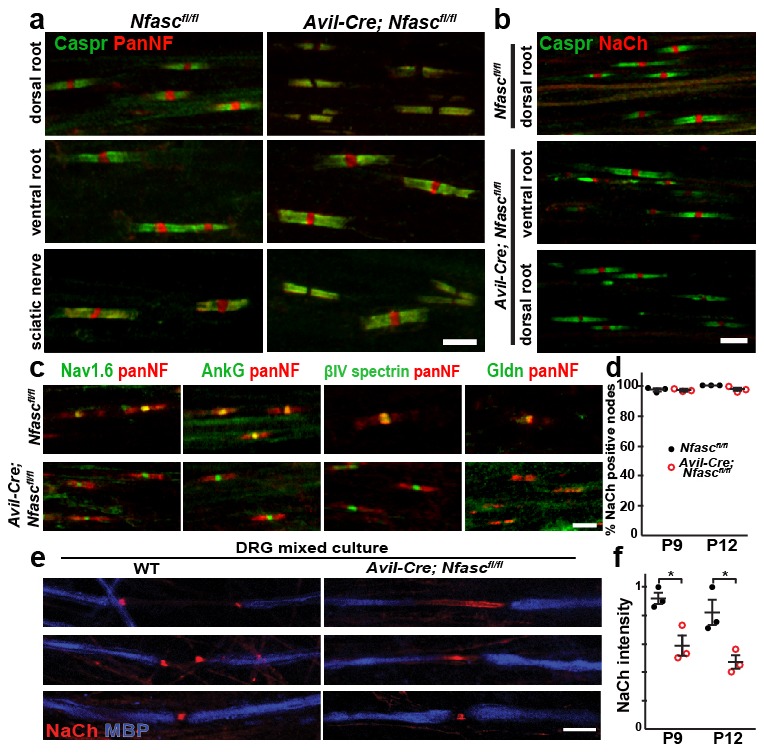
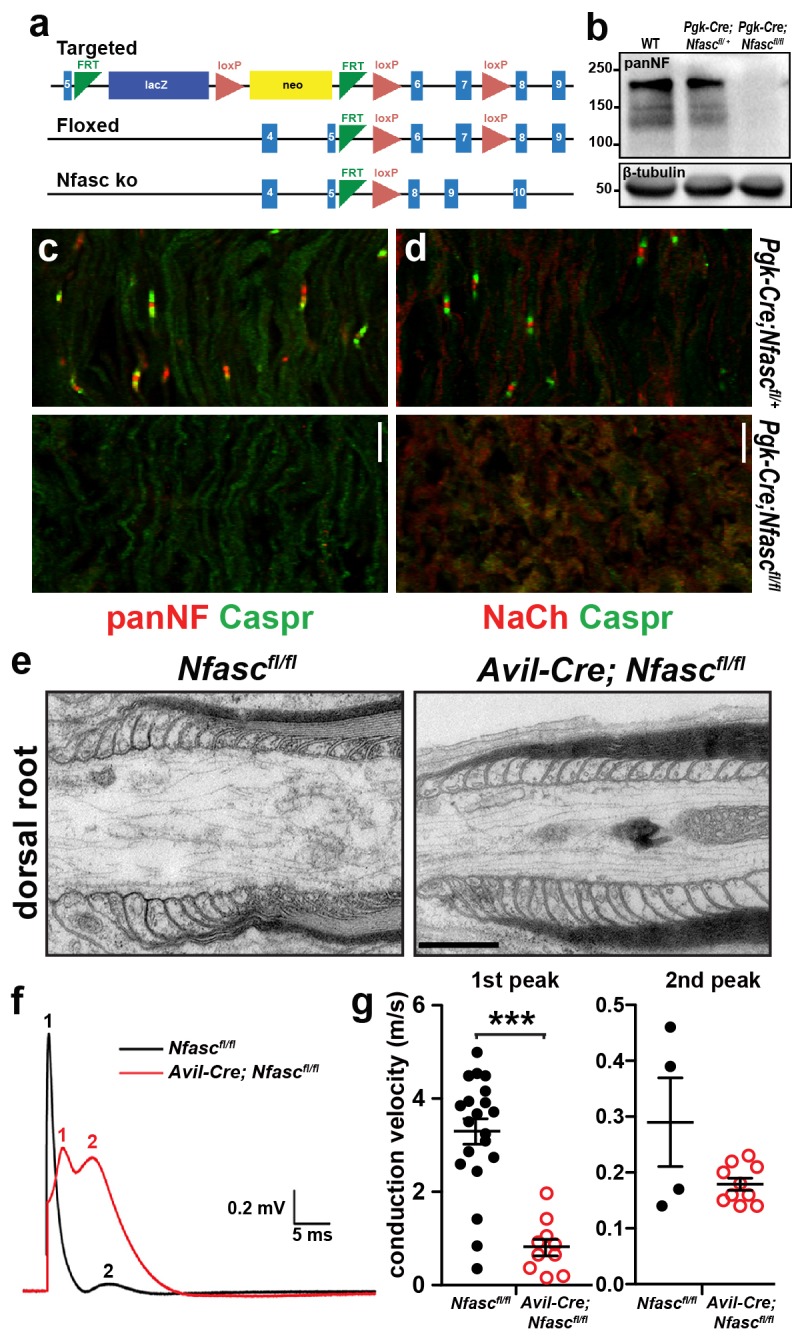
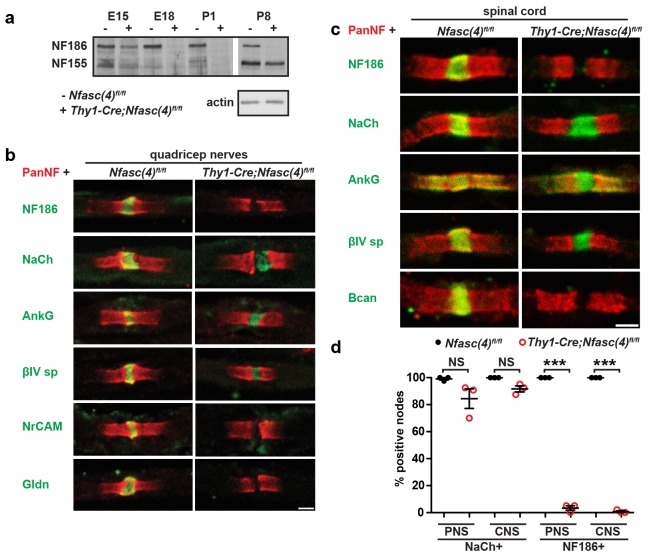
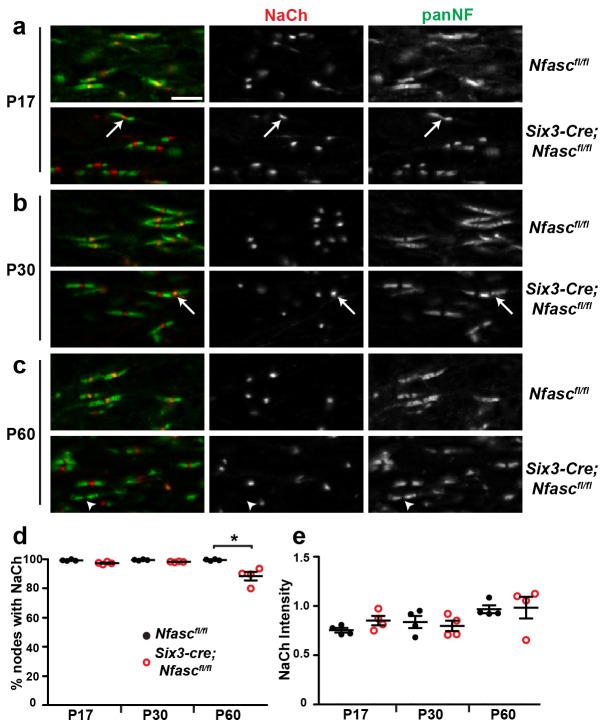
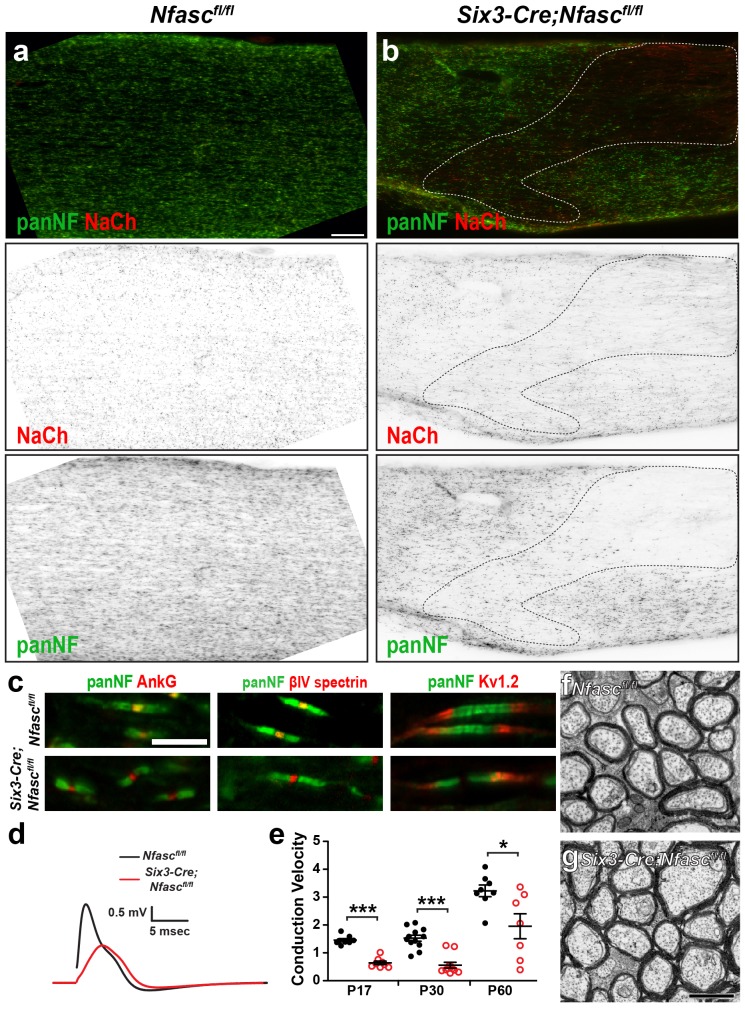
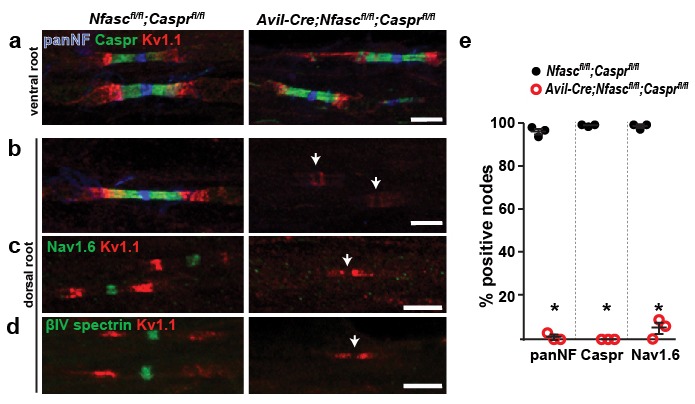
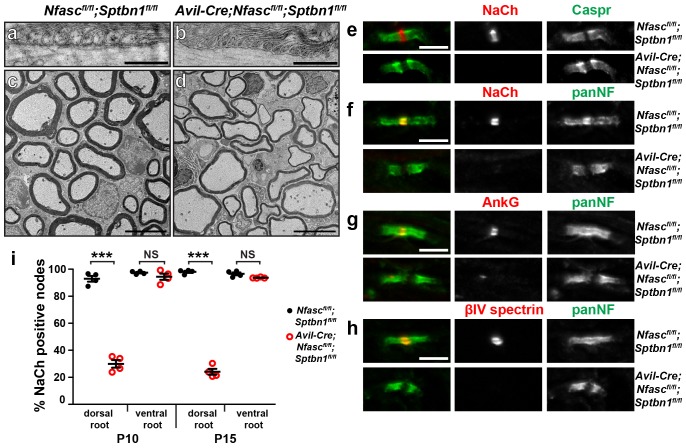
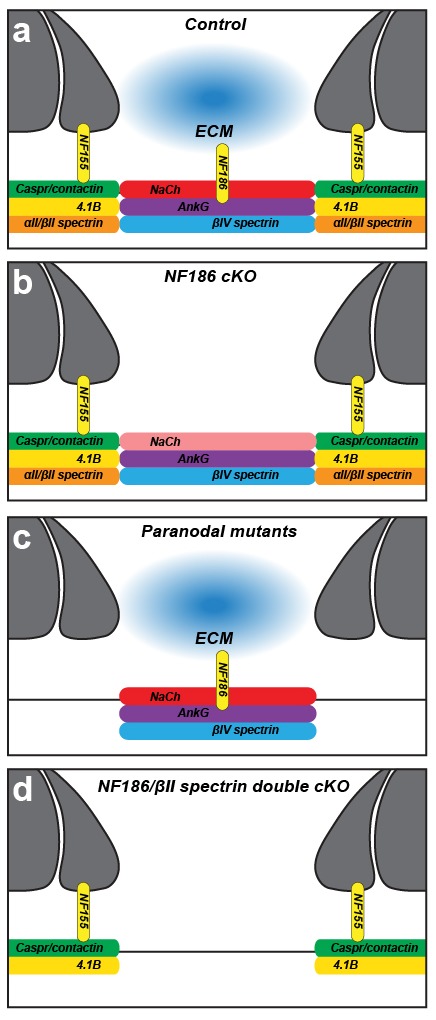
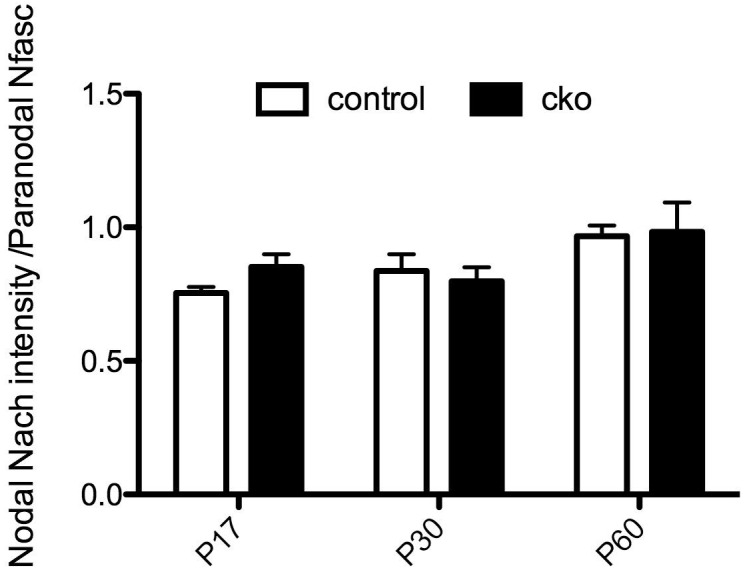
A high density of Na+ channels at nodes of Ranvier is necessary for rapid and efficient action potential propagation in myelinated axons. Na+ channel clustering is thought to depend on two axonal cell adhesion molecules that mediate interactions between the axon and myelinating glia at the nodal gap (i.e., NF186) and the paranodal junction (i.e., Caspr). Here we show that while Na+ channels cluster at nodes in the absence of NF186, they fail to do so in double conditional knockout mice lacking both NF186 and the paranodal cell adhesion molecule Caspr, demonstrating that a paranodal junction-dependent mechanism can cluster Na+ channels at nodes. Furthermore, we show that paranode-dependent clustering of nodal Na+ channels requires axonal βII spectrin which is concentrated at paranodes. Our results reveal that the paranodal junction-dependent mechanism of Na+channel clustering is mediated by the spectrin-based paranodal axonal cytoskeleton.
Keywords: axon; ion channel; mouse; myelin; neuroscience.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
References
-
- Amor V, Feinberg K, Eshed-Eisenbach Y, Vainshtein A, Frechter S, Grumet M, Rosenbluth J, Peles E. Long-term maintenance of Na+ channels at nodes of Ranvier depends on glial contact mediated by gliomedin and NrCAM. Journal of Neuroscience. 2014;34:5089–5098. doi: 10.1523/JNEUROSCI.4752-13.2014. - DOI - PMC - PubMed
-
- Bekku Y, Vargová L, Goto Y, Vorísek I, Dmytrenko L, Narasaki M, Ohtsuka A, Fässler R, Ninomiya Y, Syková E, Oohashi T. Bral1: its role in diffusion barrier formation and conduction velocity in the CNS. Journal of Neuroscience. 2010;30:3113–3123. doi: 10.1523/JNEUROSCI.5598-09.2010. - DOI - PMC - PubMed
-
- Bhat MA, Rios JC, Lu Y, Garcia-Fresco GP, Ching W, St Martin M, Li J, Einheber S, Chesler M, Rosenbluth J, Salzer JL, Bellen HJ. Axon-glia interactions and the domain organization of myelinated axons requires neurexin IV/Caspr/Paranodin. Neuron. 2001;30:369–383. doi: 10.1016/S0896-6273(01)00294-X. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
