

A central role for R7bp in the regulation of itch sensation
- PMID: 28134655
- PMCID: PMC5393936
- DOI: 10.1097/j.pain.0000000000000860
A central role for R7bp in the regulation of itch sensation
Abstract
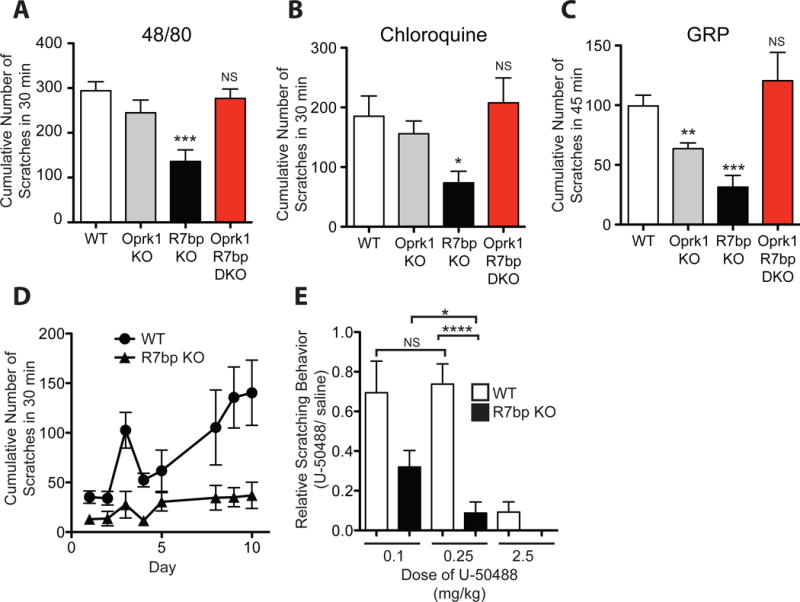
Itch is a protective sensation producing a desire to scratch. Pathologic itch can be a chronic symptom of illnesses such as uremia, cholestatic liver disease, neuropathies and dermatitis, however current therapeutic options are limited. Many types of cell surface receptors, including those present on cells in the skin, on sensory neurons and on neurons in the spinal cord, have been implicated in itch signaling. The role of G protein signaling in the regulation of pruriception is poorly understood. We identify here 2 G protein signaling components whose mutation impairs itch sensation. R7bp (a.k.a. Rgs7bp) is a palmitoylated membrane anchoring protein expressed in neurons that facilitates Gαi/o -directed GTPase activating protein activity mediated by the Gβ5/R7-RGS complex. Knockout of R7bp diminishes scratching responses to multiple cutaneously applied and intrathecally-administered pruritogens in mice. Knock-in to mice of a GTPase activating protein-insensitive mutant of Gαo (Gnao1 G184S/+) produces a similar pruriceptive phenotype. The pruriceptive defect in R7bp knockout mice was rescued in double knockout mice also lacking Oprk1, encoding the G protein-coupled kappa-opioid receptor whose activation is known to inhibit itch sensation. In a model of atopic dermatitis (eczema), R7bp knockout mice showed diminished scratching behavior and enhanced sensitivity to kappa opioid agonists. Taken together, our results indicate that R7bp is a key regulator of itch sensation and suggest the potential targeting of R7bp-dependent GTPase activating protein activity as a novel therapeutic strategy for pathological itch.
Conflict of interest statement
The authors declare no competing financial interests.
Figures






Similar articles
-
Postnatal induction and localization of R7BP, a membrane-anchoring protein for regulator of G protein signaling 7 family-Gbeta5 complexes in brain.Neuroscience. 2008 Feb 19;151(4):969-82. doi: 10.1016/j.neuroscience.2007.11.045. Epub 2007 Dec 8. Neuroscience. 2008. PMID: 18248908 Free PMC article.
-
Differential control of opioid antinociception to thermal stimuli in a knock-in mouse expressing regulator of G-protein signaling-insensitive Gαo protein.J Neurosci. 2013 Mar 6;33(10):4369-77. doi: 10.1523/JNEUROSCI.5470-12.2013. J Neurosci. 2013. PMID: 23467353 Free PMC article.
-
Regulation of neurite morphogenesis by interaction between R7 regulator of G protein signaling complexes and G protein subunit Gα13.J Biol Chem. 2017 Jun 16;292(24):9906-9918. doi: 10.1074/jbc.M116.771923. Epub 2017 Apr 21. J Biol Chem. 2017. PMID: 28432124 Free PMC article.
-
RGS-insensitive G-protein mutations to study the role of endogenous RGS proteins.Methods Enzymol. 2004;389:229-43. doi: 10.1016/S0076-6879(04)89014-1. Methods Enzymol. 2004. PMID: 15313569 Review.
-
Mas-Related G Protein-Coupled Receptors and the Biology of Itch Sensation.Annu Rev Genet. 2017 Nov 27;51:103-121. doi: 10.1146/annurev-genet-120116-024723. Annu Rev Genet. 2017. PMID: 29178819 Review.
Cited by
-
Opioidergic Signaling-A Neglected, Yet Potentially Important Player in Atopic Dermatitis.Int J Mol Sci. 2022 Apr 8;23(8):4140. doi: 10.3390/ijms23084140. Int J Mol Sci. 2022. PMID: 35456955 Free PMC article. Review.
-
Interventions in the B-type natriuretic peptide signalling pathway as a means of controlling chronic itch.Br J Pharmacol. 2020 Mar;177(5):1025-1040. doi: 10.1111/bph.14952. Epub 2020 Feb 12. Br J Pharmacol. 2020. PMID: 31877230 Free PMC article. Review.
-
Transcriptional Alterations of Mouse Trigeminal Ganglion Neurons Following Orofacial Inflammation Revealed by Single-Cell Analysis.Front Cell Neurosci. 2022 Jun 2;16:885569. doi: 10.3389/fncel.2022.885569. eCollection 2022. Front Cell Neurosci. 2022. PMID: 35722619 Free PMC article.
-
Specific regulation of mechanical nociception by Gβ5 involves GABA-B receptors.JCI Insight. 2023 Jul 10;8(13):e134685. doi: 10.1172/jci.insight.134685. JCI Insight. 2023. PMID: 37219953 Free PMC article.
-
Dynamic regulation of Rgs16 and its correlation with Neuregulin1 expression in acute and chronic nerve injury.Front Cell Dev Biol. 2025 Mar 27;13:1540453. doi: 10.3389/fcell.2025.1540453. eCollection 2025. Front Cell Dev Biol. 2025. PMID: 40213396 Free PMC article.
References
-
- Anderson GR, Lujan R, Semenov A, Pravetoni M, Posokhova EN, Song JH, Uversky V, Chen CK, Wickman K, Martemyanov KA. Expression and localization of RGS9-2/G 5/R7BP complex in vivo is set by dynamic control of its constitutive degradation by cellular cysteine proteases. J Neurosci. 2007;27(51):14117–14127. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
