

Hormonal gain control of a medial preoptic area social reward circuit
- PMID: 28135243
- PMCID: PMC5735833
- DOI: 10.1038/nn.4487
Hormonal gain control of a medial preoptic area social reward circuit
Erratum in
-
Corrigendum: Hormonal gain control of a medial preoptic area social reward circuit.Nat Neurosci. 2017 Sep 28;20(10):1427-1430. doi: 10.1038/nn1017-1427c. Nat Neurosci. 2017. PMID: 28949330 No abstract available.
Abstract
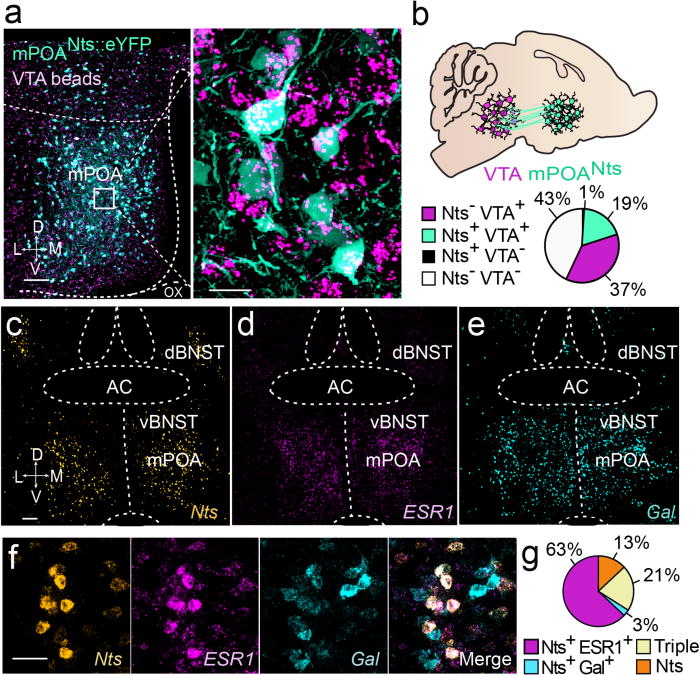
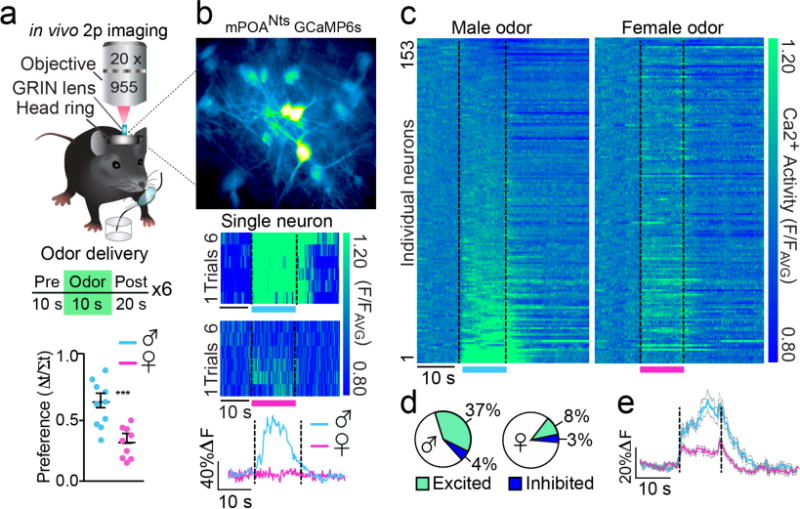
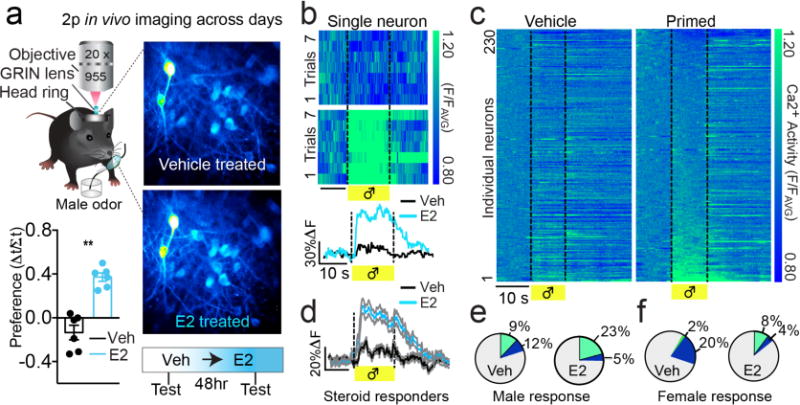
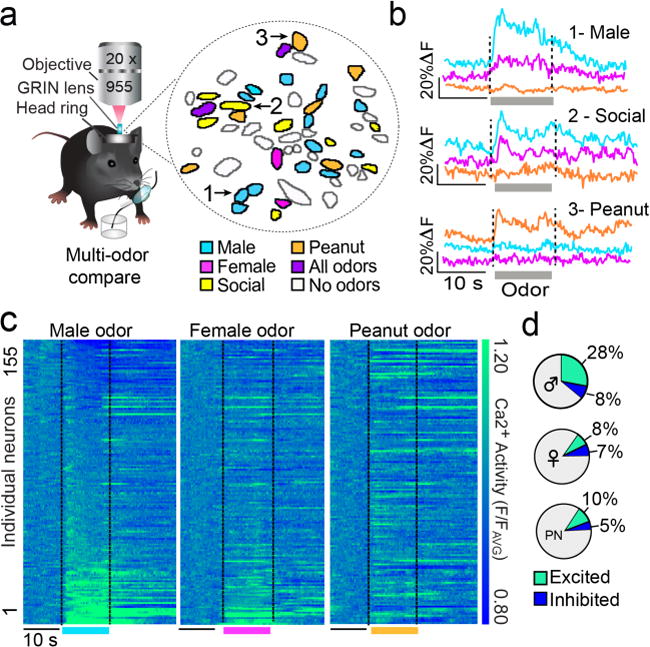
Neural networks that control reproduction must integrate social and hormonal signals, tune motivation, and coordinate social interactions. However, the neural circuit mechanisms for these processes remain unresolved. The medial preoptic area (mPOA), an essential node for social behaviors, comprises molecularly diverse neurons with widespread projections. Here we identify a steroid-responsive subset of neurotensin (Nts)-expressing mPOA neurons that interface with the ventral tegmental area (VTA) to form a socially engaged reward circuit. Using in vivo two-photon imaging in female mice, we show that mPOANts neurons preferentially encode attractive male cues compared to nonsocial appetitive stimuli. Ovarian hormone signals regulate both the physiological and cue-encoding properties of these cells. Furthermore, optogenetic stimulation of mPOANts-VTA circuitry promotes rewarding phenotypes, social approach and striatal dopamine release. Collectively, these data demonstrate that steroid-sensitive mPOA neurons encode ethologically relevant stimuli and co-opt midbrain reward circuits to promote prosocial behaviors critical for species survival.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
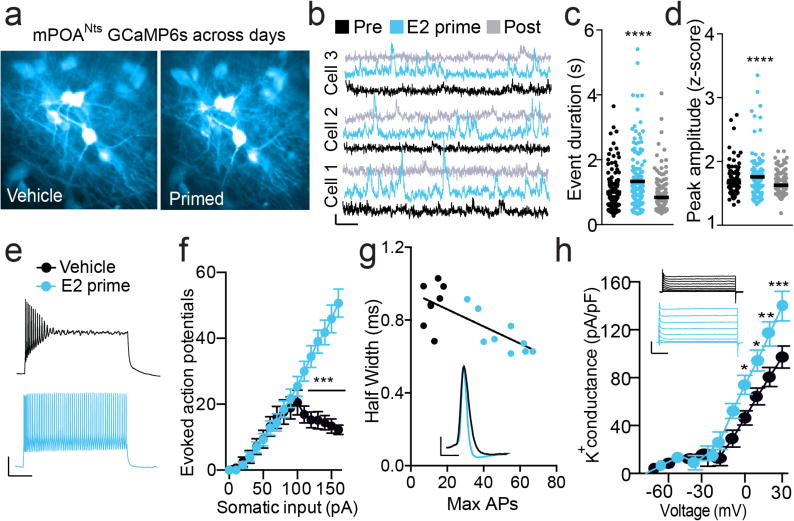
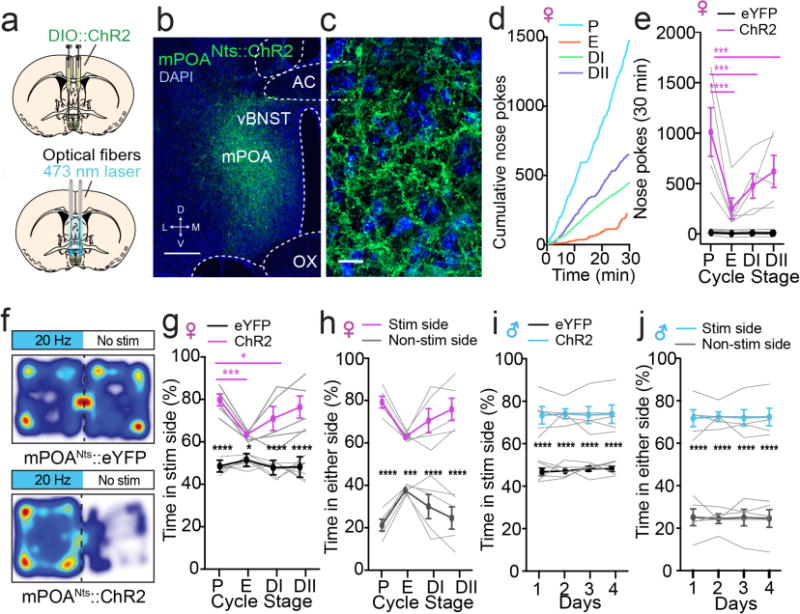
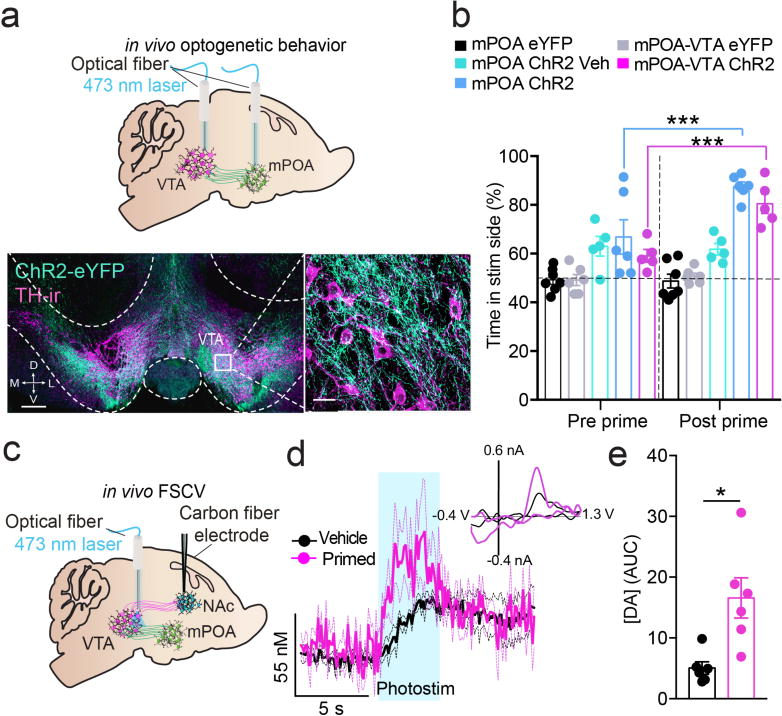
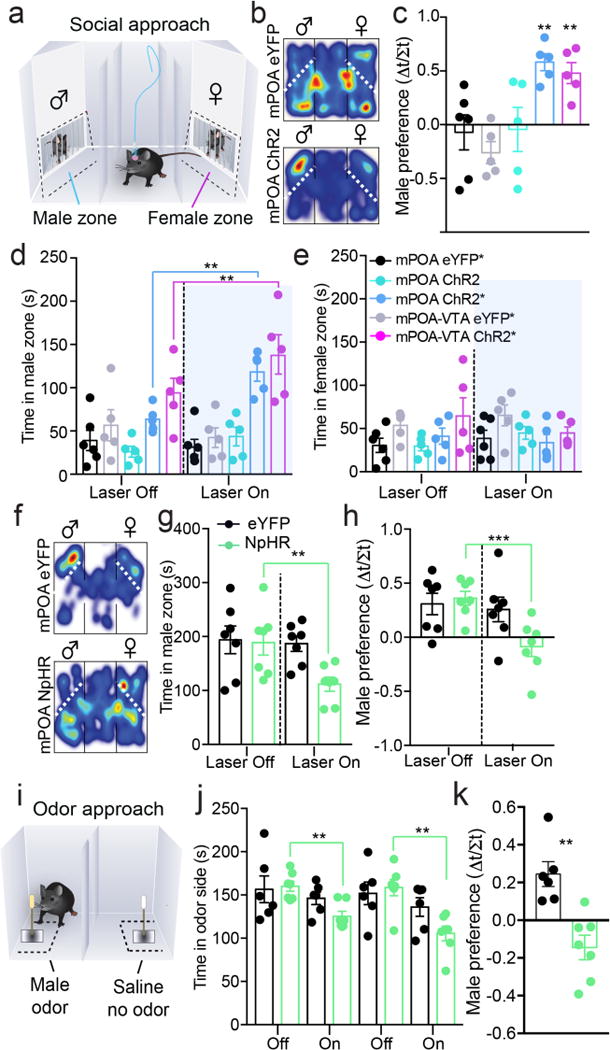
Comment in
-
Addicted to love?Sci Transl Med. 2017 Feb 22;9(378):eaam6067. doi: 10.1126/scitranslmed.aam6067. Sci Transl Med. 2017. PMID: 28228599
-
Setting the mood for love.Nat Neurosci. 2017 Feb 23;20(3):379-380. doi: 10.1038/nn.4514. Nat Neurosci. 2017. PMID: 28230842 No abstract available.
References
Online Methods References
-
- Edwards DA. Induction of estrus in female mice: Estrogen-progesterone interactions. Hormones and Behavior. 1970;1:299–304.
MeSH terms
Substances
Grants and funding
- K01 AA023555/AA/NIAAA NIH HHS/United States
- U01 AA020911/AA/NIAAA NIH HHS/United States
- F30 AA021312/AA/NIAAA NIH HHS/United States
- P30 NS045892/NS/NINDS NIH HHS/United States
- R01 AA022449/AA/NIAAA NIH HHS/United States
- F32 DA041184/DA/NIDA NIH HHS/United States
- T32 NS007431/NS/NINDS NIH HHS/United States
- T32 MH093315/MH/NIMH NIH HHS/United States
- F32 DK112564/DK/NIDDK NIH HHS/United States
- R01 DA038168/DA/NIDA NIH HHS/United States
- R01 DA032750/DA/NIDA NIH HHS/United States
- R37 DA032750/DA/NIDA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
