

Cellular Immune Responses to Live Attenuated Japanese Encephalitis (JE) Vaccine SA14-14-2 in Adults in a JE/Dengue Co-Endemic Area
- PMID: 28135273
- PMCID: PMC5279729
- DOI: 10.1371/journal.pntd.0005263
Cellular Immune Responses to Live Attenuated Japanese Encephalitis (JE) Vaccine SA14-14-2 in Adults in a JE/Dengue Co-Endemic Area
Abstract
Background: Japanese encephalitis (JE) virus (JEV) causes severe epidemic encephalitis across Asia, for which the live attenuated vaccine SA14-14-2 is being used increasingly. JEV is a flavivirus, and is closely related to dengue virus (DENV), which is co-endemic in many parts of Asia, with clinically relevant interactions. There is no information on the human T cell response to SA14-14-2, or whether responses to SA14-14-2 cross-react with DENV. We used live attenuated JE vaccine SA14-14-2 as a model for studying T cell responses to JEV infection in adults, and to determine whether these T cell responses are cross-reactive with DENV, and other flaviviruses.
Methods: We conducted a single arm, open label clinical trial (registration: clinicaltrials.gov NCT01656200) to study T cell responses to SA14-14-2 in adults in South India, an area endemic for JE and dengue.
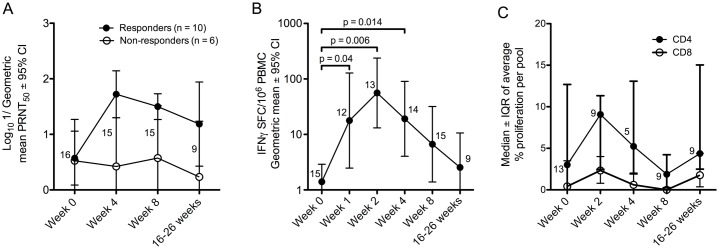
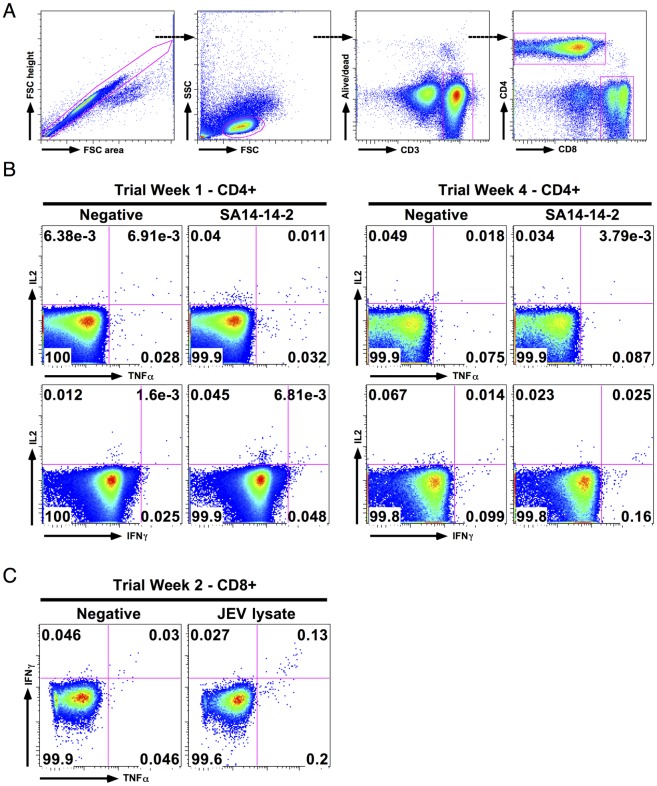
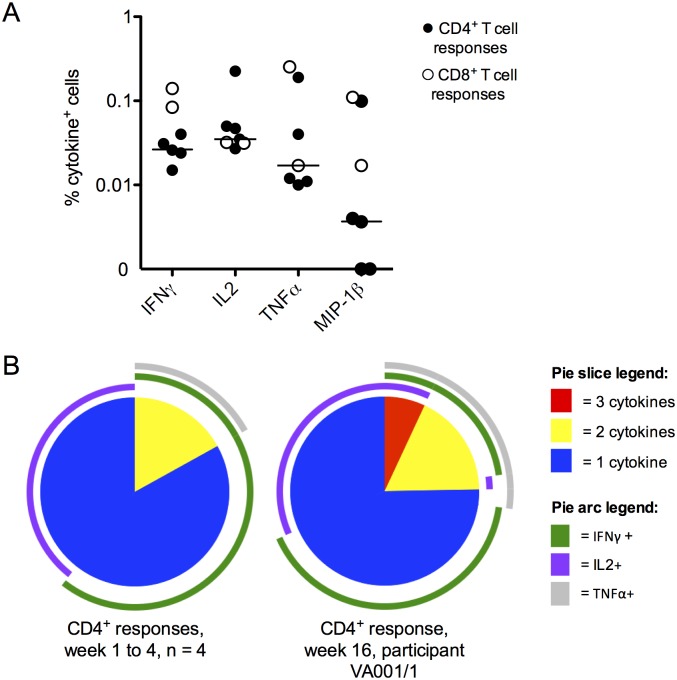
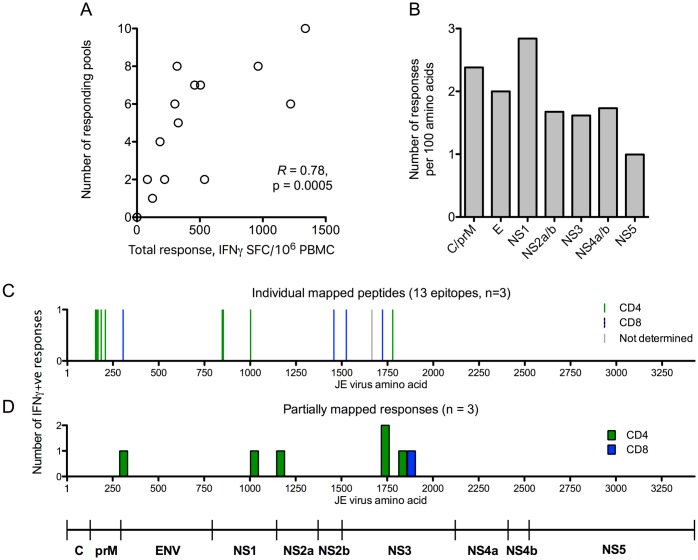
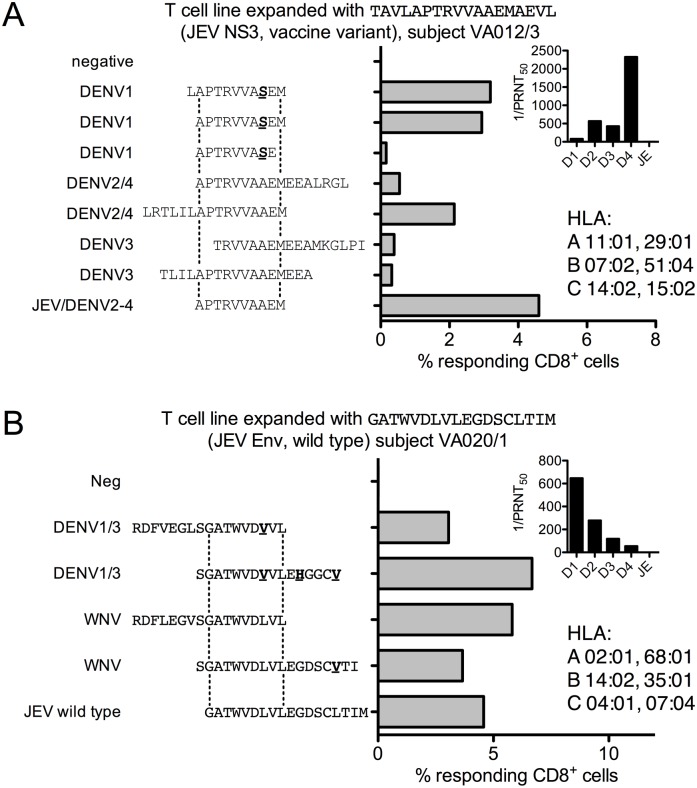
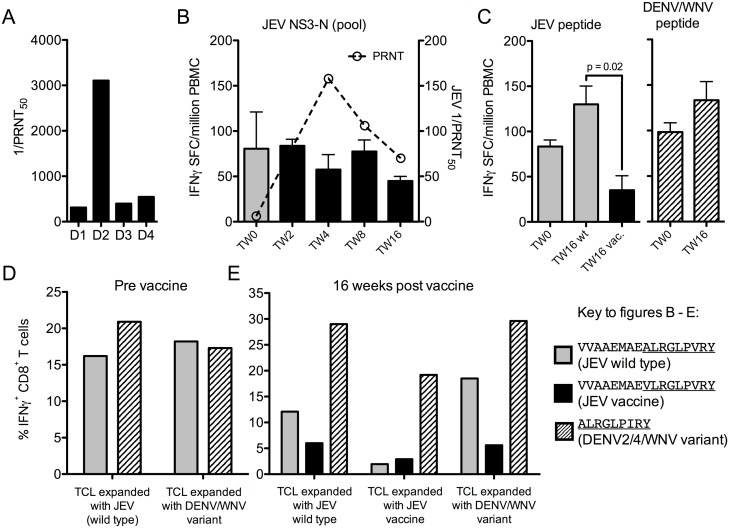
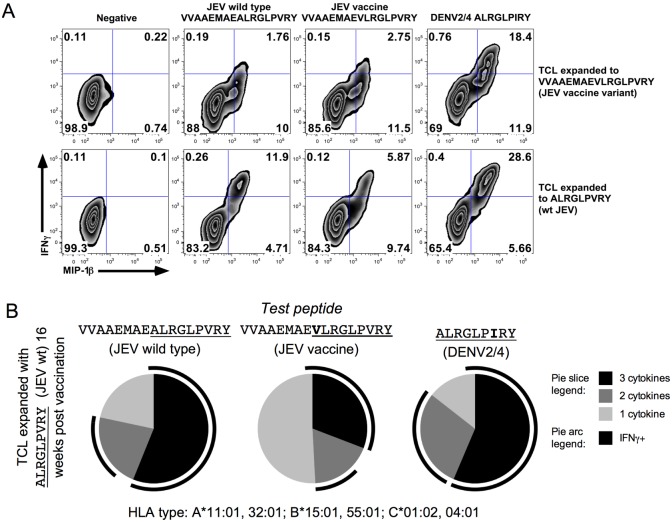
Results: Ten out of 16 (62.5%) participants seroconverted to JEV SA14-14-2, and geometric mean neutralising antibody (NAb) titre was 18.5. Proliferation responses were commonly present before vaccination in the absence of NAb, indicating a likely high degree of previous flavivirus exposure. Thirteen of 15 (87%) participants made T cell interferon-gamma (IFNγ) responses against JEV proteins. In four subjects tested, at least some T cell epitopes mapped cross-reacted with DENV and other flaviviruses.
Conclusions: JEV SA14-14-2 was more immunogenic for T cell IFNγ than for NAb in adults in this JE/DENV co-endemic area. The proliferation positive, NAb negative combination may represent a new marker of long term immunity/exposure to JE. T cell responses can cross-react between JE vaccine and DENV in a co-endemic area, illustrating a need for greater knowledge on such responses to inform the development of next-generation vaccines effective against both diseases.
Trial registration: clinicaltrials.gov (NCT01656200).
Conflict of interest statement
MY is an employee of Vaxyn Consulting. Vaxyn Consulting has no financial interest in JE vaccine SA14-14-2. The other authors have declared that no competing interests exist.
Figures
References
-
- Sumiyoshi H, Mori C, Fuke I, Morita K, Kuhara S, Kondou J, et al. Complete nucleotide sequence of the Japanese encephalitis virus genome RNA. Virology. 1987;161(2):497–510. - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
