

AMH/MIS as a contraceptive that protects the ovarian reserve during chemotherapy
- PMID: 28137855
- PMCID: PMC5338508
- DOI: 10.1073/pnas.1620729114
AMH/MIS as a contraceptive that protects the ovarian reserve during chemotherapy
Abstract
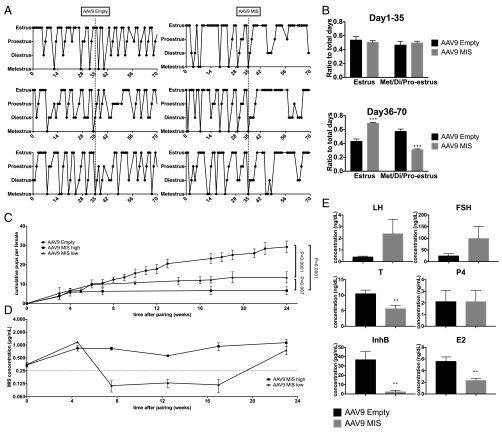
The ovarian reserve represents the stock of quiescent primordial follicles in the ovary which is gradually depleted during a woman's reproductive lifespan, resulting in menopause. Müllerian inhibiting substance (MIS) (or anti-Müllerian hormone/AMH), which is produced by granulosa cells of growing follicles, has been proposed as a negative regulator of primordial follicle activation. Here we show that long-term parenteral administration of superphysiological doses of MIS, using either an adeno-associated virus serotype 9 (AAV9) gene therapy vector or recombinant protein, resulted in a complete arrest of folliculogenesis in mice. The ovaries of MIS-treated mice were smaller than those in controls and did not contain growing follicles but retained a normal ovarian reserve. When mice treated with AAV9/MIS were paired with male breeders, they exhibited complete and permanent contraception for their entire reproductive lifespan, disrupted vaginal cycling, and hypergonadotropic hypogonadism. However, when ovaries from AAV9-MIS-treated mice were transplanted orthotopically into normal recipient mice, or when treatment with the protein was discontinued, folliculogenesis resumed, suggesting reversibility. One of the important causes of primary ovarian insufficiency is chemotherapy-induced primordial follicle depletion, which has been proposed to be mediated in part by increased activation. To test the hypothesis that MIS could prevent chemotherapy-induced overactivation, mice were given carboplatin, doxorubicin, or cyclophosphamide and were cotreated with AAV9-MIS, recombinant MIS protein, or vehicle controls. We found significantly more primordial follicles in MIS-treated animals than in controls. Thus treatment with MIS may provide a method of contraception with the unique characteristic of blocking primordial follicle activation that could be exploited to prevent the primary ovarian insufficiency often associated with chemotherapy.
Keywords: AAV9; AMH; MIS; contraceptive; oncofertility.
Conflict of interest statement
The authors declare no conflict of interest.
Figures








Comment in
-
A win-win for women's reproductive health: A nonsteroidal contraceptive and fertoprotective neoadjuvant.Proc Natl Acad Sci U S A. 2017 Feb 28;114(9):2101-2102. doi: 10.1073/pnas.1700337114. Epub 2017 Feb 17. Proc Natl Acad Sci U S A. 2017. PMID: 28213496 Free PMC article. No abstract available.
References
-
- Baker ML, Metcalfe SA, Hutson JM. Serum levels of Müllerian inhibiting substance in boys from birth to 18 years, as determined by enzyme immunoassay. J Clin Endocrinol Metab. 1990;70(1):11–15. - PubMed
-
- Hudson PL, et al. An immunoassay to detect human müllerian inhibiting substance in males and females during normal development. J Clin Endocrinol Metab. 1990;70(1):16–22. - PubMed
-
- Josso N, Legeai L, Forest MG, Chaussain J-L, Brauner R. An enzyme linked immunoassay for anti-müllerian hormone: A new tool for the evaluation of testicular function in infants and children. J Clin Endocrinol Metab. 1990;70(1):23–27. - PubMed
-
- Seifer DB, et al. Gonadotropin-releasing hormone agonist-induced differences in granulosa cell cycle kinetics are associated with alterations in follicular fluid müllerian-inhibiting substance and androgen content. J Clin Endocrinol Metab. 1993;76(3):711–714. - PubMed
-
- Josso N, Picard JY, Rey R, di Clemente N. Testicular anti-Müllerian hormone: History, genetics, regulation and clinical applications. Pediatr Endocrinol Rev. 2006;3(4):347–358. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
