

Many roads to symmetry breaking: molecular mechanisms and theoretical models of yeast cell polarity
- PMID: 28137950
- PMCID: PMC5341721
- DOI: 10.1091/mbc.E16-10-0739
Many roads to symmetry breaking: molecular mechanisms and theoretical models of yeast cell polarity
Abstract
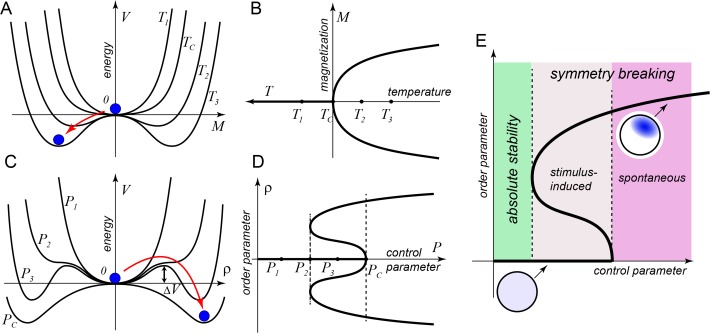
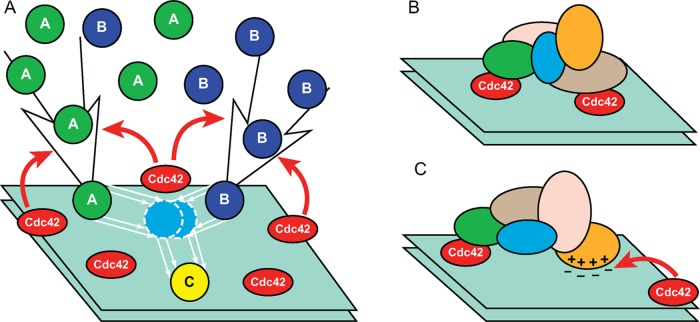
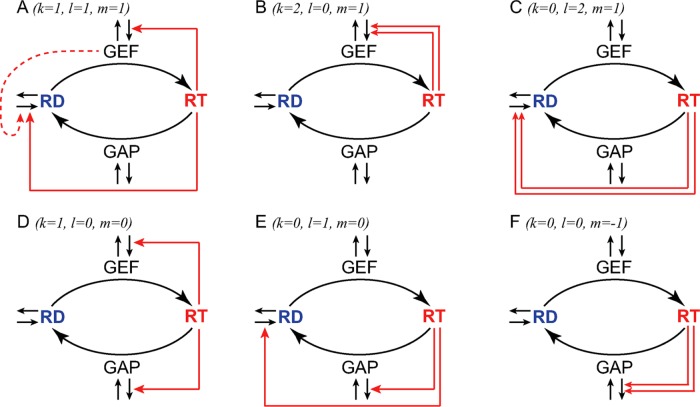
Mathematical modeling has been instrumental in identifying common principles of cell polarity across diverse systems. These principles include positive feedback loops that are required to destabilize a spatially uniform state of the cell. The conserved small G-protein Cdc42 is a master regulator of eukaryotic cellular polarization. Here we discuss recent developments in studies of Cdc42 polarization in budding and fission yeasts and demonstrate that models describing symmetry-breaking polarization can be classified into six minimal classes based on the structure of positive feedback loops that activate and localize Cdc42. Owing to their generic system-independent nature, these model classes are also likely to be relevant for the G-protein-based symmetry-breaking systems of higher eukaryotes. We review experimental evidence pro et contra different theoretically plausible models and conclude that several parallel and non-mutually exclusive mechanisms are likely involved in cellular polarization of yeasts. This potential redundancy needs to be taken into consideration when interpreting the results of recent cell-rewiring studies.
© 2017 Goryachev and Leda. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures
References
-
- Atkins BD, Yoshida S, Pellman D. Symmetry breaking: scaffold plays matchmaker for polarity signaling proteins. Curr Biol. 2008;18:R1130–R1132. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
