

Serotonin-dependent kinetics of feeding bursts underlie a graded response to food availability in C. elegans
- PMID: 28145493
- PMCID: PMC5296638
- DOI: 10.1038/ncomms14221
Serotonin-dependent kinetics of feeding bursts underlie a graded response to food availability in C. elegans
Abstract
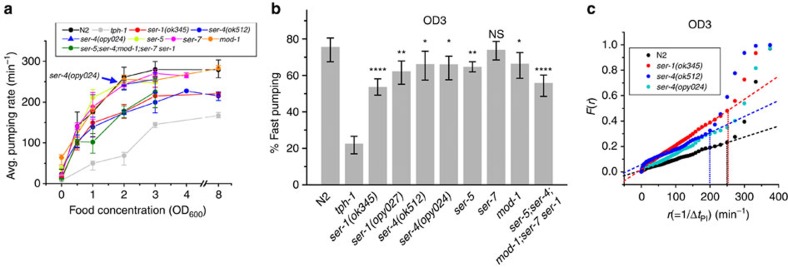
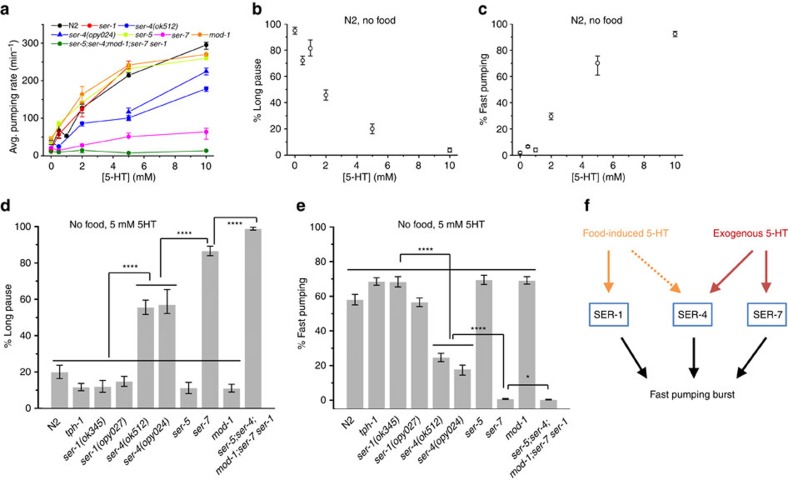
Animals integrate physiological and environmental signals to modulate their food uptake. The nematode C. elegans, whose food uptake consists of pumping bacteria from the environment into the gut, provides excellent opportunities for discovering principles of conserved regulatory mechanisms. Here we show that worms implement a graded feeding response to the concentration of environmental bacteria by modulating a commitment to bursts of fast pumping. Using long-term, high-resolution, longitudinal recordings of feeding dynamics under defined conditions, we find that the frequency and duration of pumping bursts increase and the duration of long pauses diminishes in environments richer in bacteria. The bioamine serotonin is required for food-dependent induction of bursts as well as for maintaining their high rate of pumping through two distinct mechanisms. We identify the differential roles of distinct families of serotonin receptors in this process and propose that regulation of bursts is a conserved mechanism of behaviour and motor control.
Conflict of interest statement
The authors declare no competing financial interests.
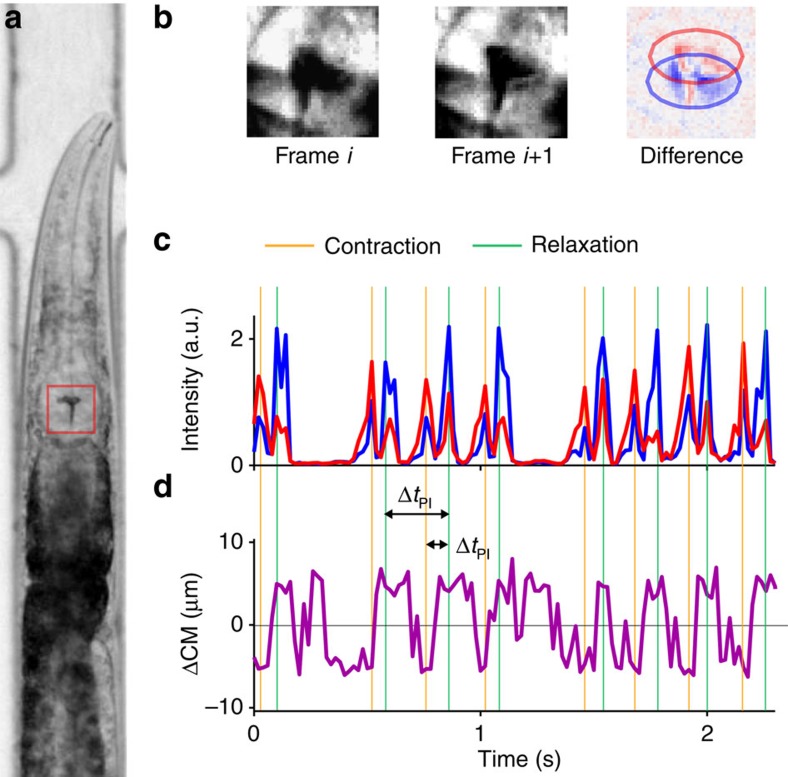
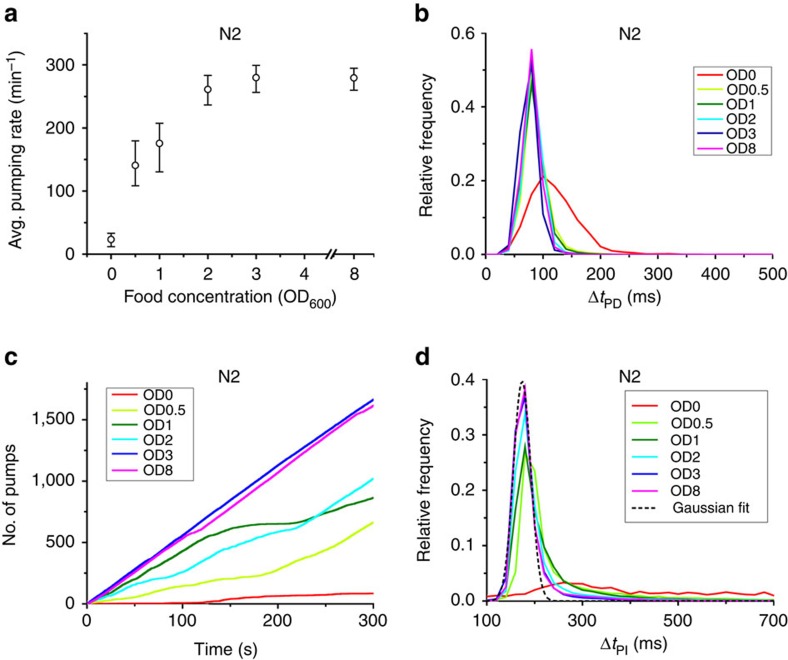
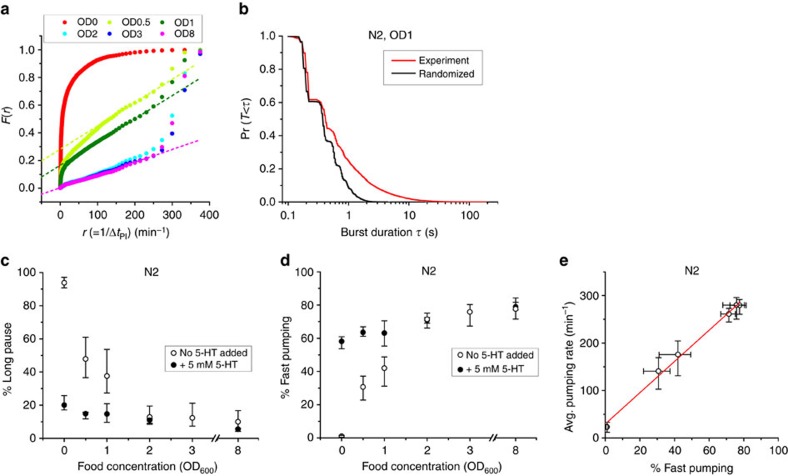
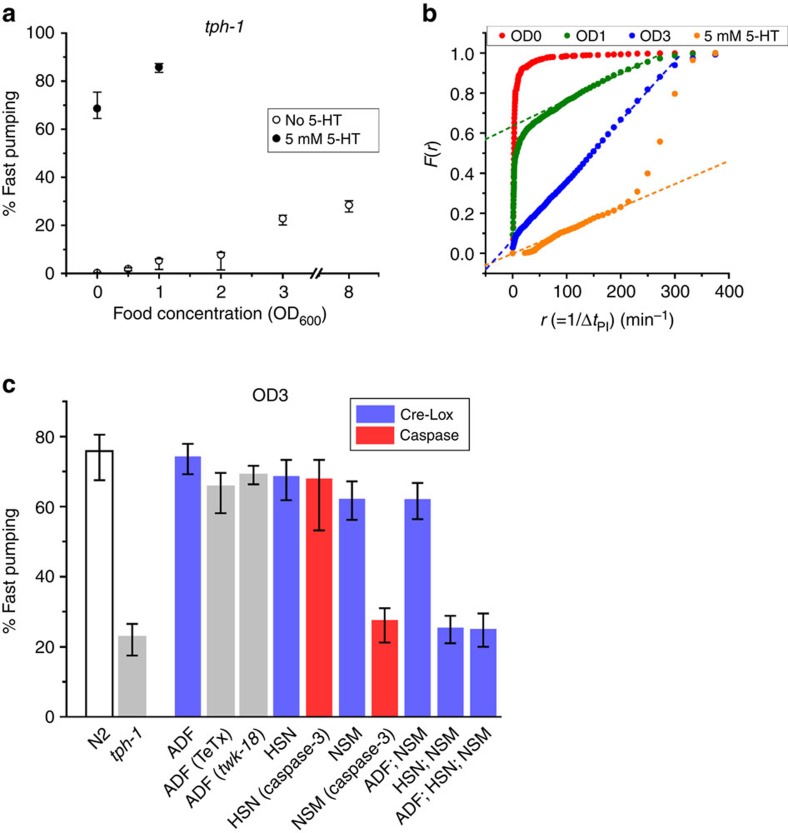
Figures
References
-
- Berthoud H. R. & Morrison C. The brain, appetite, and obesity. Annu. Rev. Psychol. 59, 55–92 (2008). - PubMed
-
- Halford J. C., Boyland E. J., Blundell J. E., Kirkham T. C. & Harrold J. A. Pharmacological management of appetite expression in obesity. Nat. Rev. Endocrinol. 6, 255–269 (2010). - PubMed
-
- Tecott L. H. et al.. Eating disorder and epilepsy in mice lacking 5-HT2c serotonin receptors. Nature 374, 542–546 (1995). - PubMed
-
- Kenyon C. J. The genetics of ageing. Nature 464, 504–512 (2010). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
