

Genome-wide exonic small interference RNA-mediated gene silencing regulates sexual reproduction in the homothallic fungus Fusarium graminearum
- PMID: 28146558
- PMCID: PMC5310905
- DOI: 10.1371/journal.pgen.1006595
Genome-wide exonic small interference RNA-mediated gene silencing regulates sexual reproduction in the homothallic fungus Fusarium graminearum
Abstract
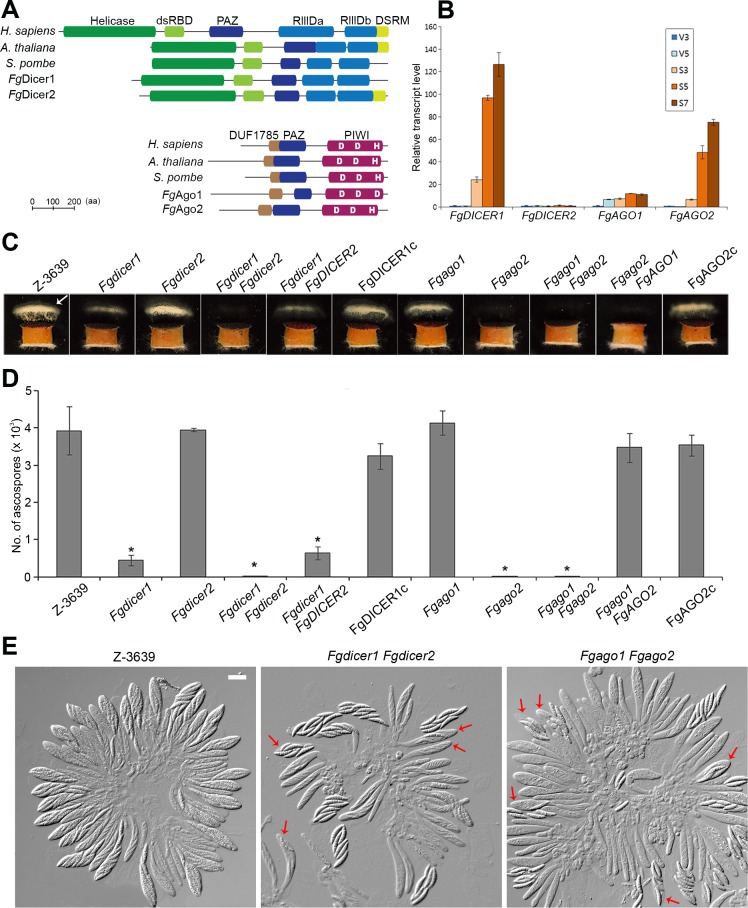
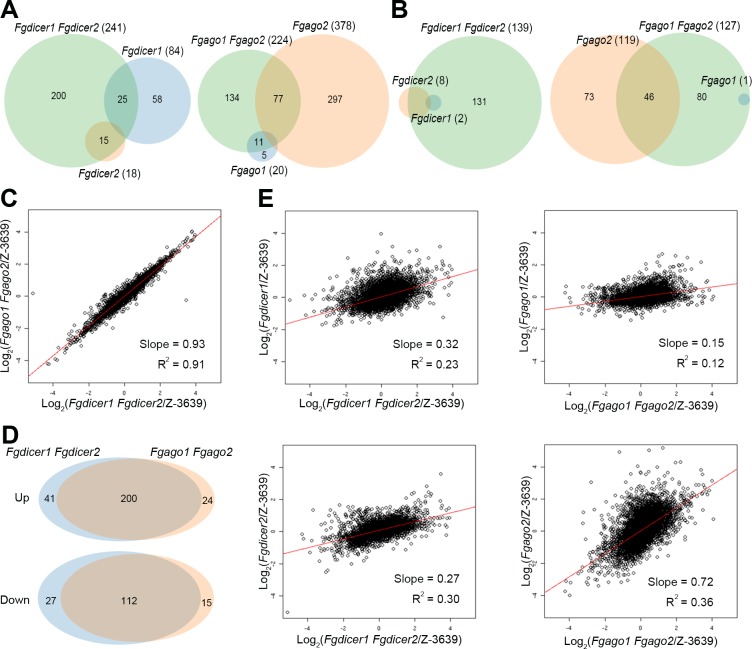
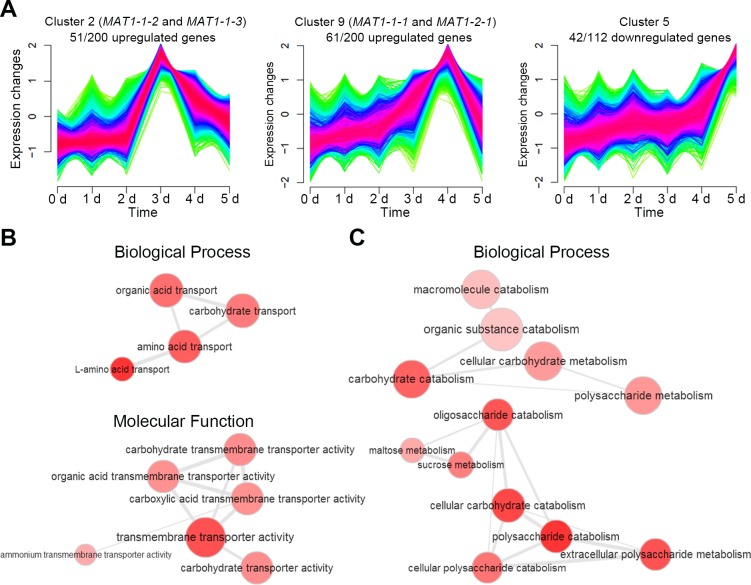
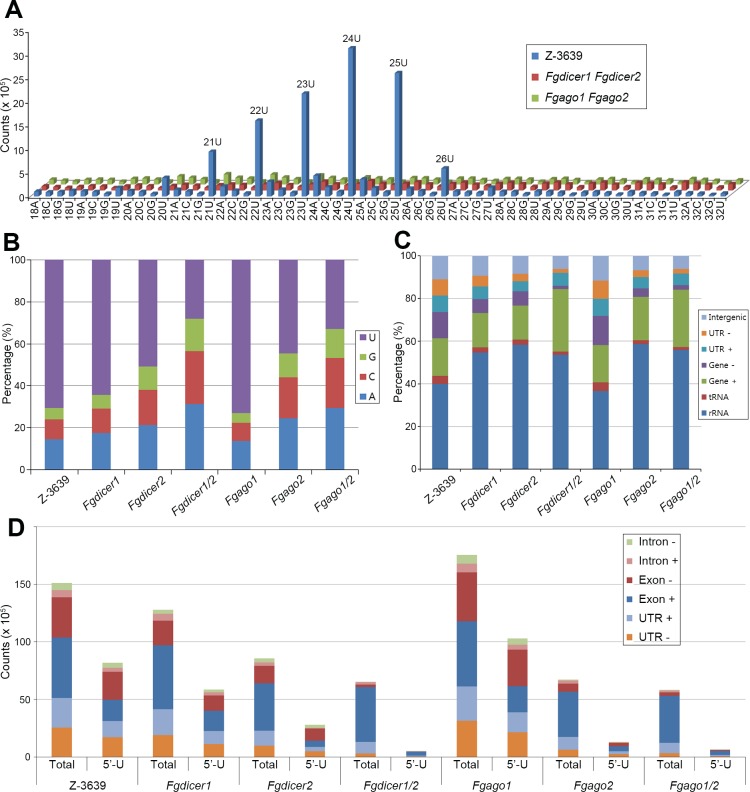
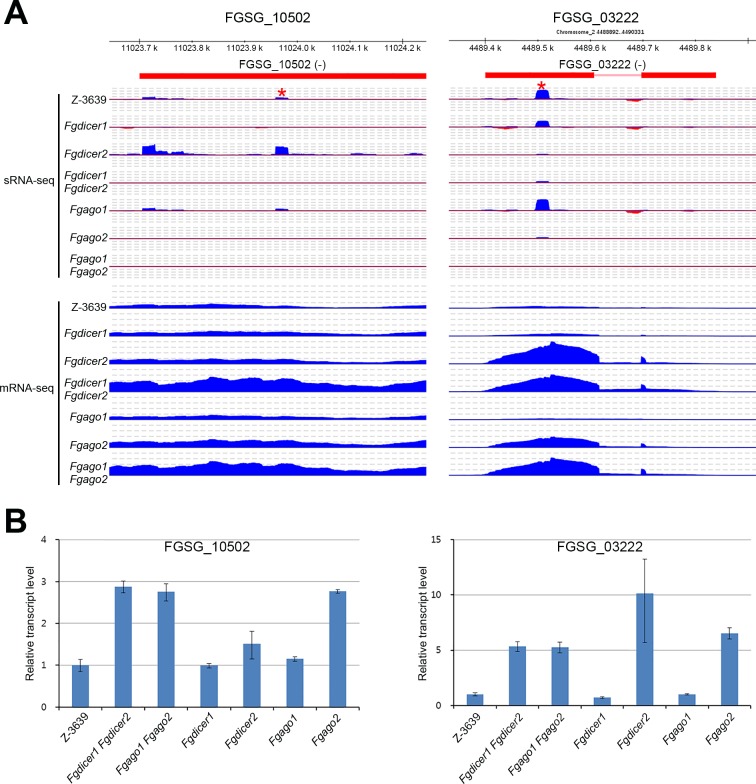
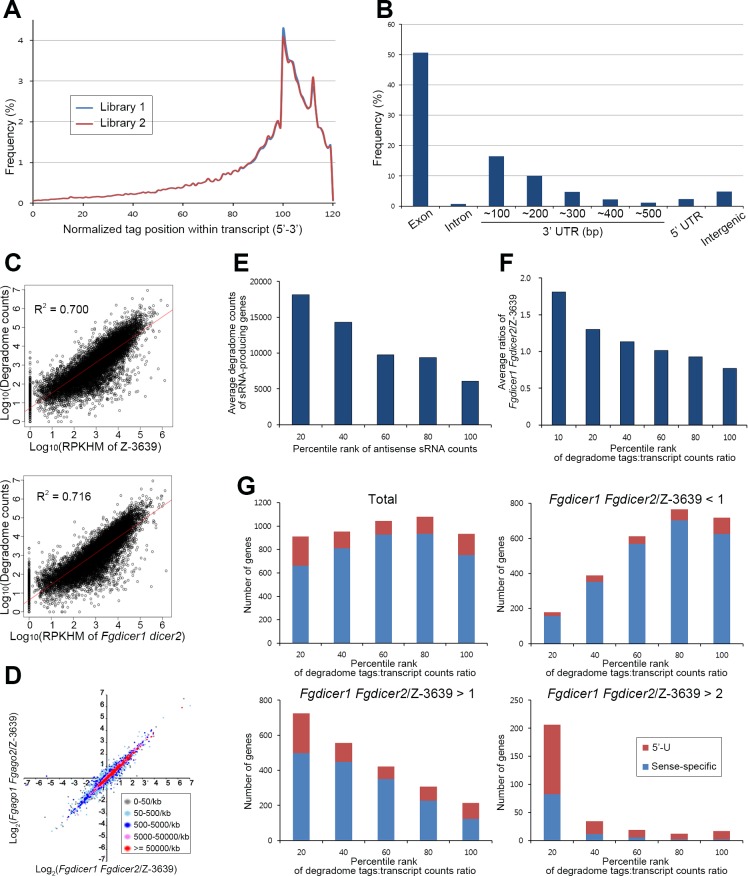
Various ascomycete fungi possess sex-specific molecular mechanisms, such as repeat-induced point mutations, meiotic silencing by unpaired DNA, and unusual adenosine-to-inosine RNA editing, for genome defense or gene regulation. Using a combined analysis of functional genetics and deep sequencing of small noncoding RNA (sRNA), mRNA, and the degradome, we found that the sex-specifically induced exonic small interference RNA (ex-siRNA)-mediated RNA interference (RNAi) mechanism has an important role in fine-tuning the transcriptome during ascospore formation in the head blight fungus Fusarium graminearum. Approximately one-third of the total sRNAs were produced from the gene region, and sRNAs with an antisense direction or 5'-U were involved in post-transcriptional gene regulation by reducing the stability of the corresponding gene transcripts. Although both Dicers and Argonautes partially share their functions, the sex-specific RNAi pathway is primarily mediated by FgDicer1 and FgAgo2, while the constitutively expressed RNAi components FgDicer2 and FgAgo1 are responsible for hairpin-induced RNAi. Based on our results, we concluded that F. graminearum primarily utilizes ex-siRNA-mediated RNAi for ascosporogenesis but not for genome defenses and other developmental stages. Each fungal species appears to have evolved RNAi-based gene regulation for specific developmental stages or stress responses. This study provides new insights into the regulatory role of sRNAs in fungi and other lower eukaryotes.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


Similar articles
-
Characterization of RNA silencing components in the plant pathogenic fungus Fusarium graminearum.Sci Rep. 2015 Jul 27;5:12500. doi: 10.1038/srep12500. Sci Rep. 2015. PMID: 26212591 Free PMC article.
-
RNA-dependent RNA polymerases regulate ascospore discharge through the exonic-sRNA-mediated RNAi pathway.mBio. 2024 Jun 12;15(6):e0037724. doi: 10.1128/mbio.00377-24. Epub 2024 May 16. mBio. 2024. PMID: 38752738 Free PMC article.
-
Different Components of the RNA Interference Machinery Are Required for Conidiation, Ascosporogenesis, Virulence, Deoxynivalenol Production, and Fungal Inhibition by Exogenous Double-Stranded RNA in the Head Blight Pathogen Fusarium graminearum.Front Microbiol. 2019 Aug 7;10:1662. doi: 10.3389/fmicb.2019.01662. eCollection 2019. Front Microbiol. 2019. PMID: 31616385 Free PMC article.
-
RNA Interference in Fungi: Retention and Loss.Microbiol Spectr. 2016 Dec;4(6). doi: 10.1128/microbiolspec.FUNK-0008-2016. Microbiol Spectr. 2016. PMID: 28087943 Review.
-
RNA interference pathways in fungi: mechanisms and functions.Annu Rev Microbiol. 2012;66:305-23. doi: 10.1146/annurev-micro-092611-150138. Epub 2012 Jun 28. Annu Rev Microbiol. 2012. PMID: 22746336 Free PMC article. Review.
Cited by
-
RNAi-Based Approaches to Control Mycotoxin Producers: Challenges and Perspectives.J Fungi (Basel). 2024 Sep 29;10(10):682. doi: 10.3390/jof10100682. J Fungi (Basel). 2024. PMID: 39452634 Free PMC article. Review.
-
Host-Induced Silencing of Fusarium graminearum Genes Enhances the Resistance of Brachypodium distachyon to Fusarium Head Blight.Front Plant Sci. 2019 Oct 30;10:1362. doi: 10.3389/fpls.2019.01362. eCollection 2019. Front Plant Sci. 2019. PMID: 31737001 Free PMC article.
-
Fusarium graminearum DICER-like-dependent sRNAs are required for the suppression of host immune genes and full virulence.PLoS One. 2021 Aug 5;16(8):e0252365. doi: 10.1371/journal.pone.0252365. eCollection 2021. PLoS One. 2021. PMID: 34351929 Free PMC article.
-
Omics Approaches for Understanding Biogenesis, Composition and Functions of Fungal Extracellular Vesicles.Front Genet. 2021 May 3;12:648524. doi: 10.3389/fgene.2021.648524. eCollection 2021. Front Genet. 2021. PMID: 34012462 Free PMC article. Review.
-
Trichoderma atroviride hyphal regeneration and conidiation depend on cell-signaling processes regulated by a microRNA-like RNA.Microb Genom. 2022 Oct;8(10):mgen000869. doi: 10.1099/mgen.0.000869. Microb Genom. 2022. PMID: 36239595 Free PMC article.
References
-
- Leslie JF, Summerell BA. The Fusarium laboratory manual. Ames, Iowa: Blackwell Publisher; 2006.
-
- Maldonado-Ramirez SL, Schmale DG III, Shields EJ, Bergstrom GC. The relative abundance of viable spores of Gibberella zeae in the planetary boundary layer suggests the role of long-distance transport in regional epidemics of Fusarium head blight. Agr Forest Meteorol. 2005;132: 20–27.
-
- Dill-Macky R, Jones RK. The effect of previous crop residues and tillage on Fusarium head blight of wheat. Plant Dis. 2000;84: 71–76. - PubMed
-
- Trail F, Common R. Perithecial development by Gibberella zeae: a light microscopy study. Mycologia. 2000;92: 130–138.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
