

Natural Selection in Cancer Biology: From Molecular Snowflakes to Trait Hallmarks
- PMID: 28148564
- PMCID: PMC5287060
- DOI: 10.1101/cshperspect.a029652
Natural Selection in Cancer Biology: From Molecular Snowflakes to Trait Hallmarks
Abstract
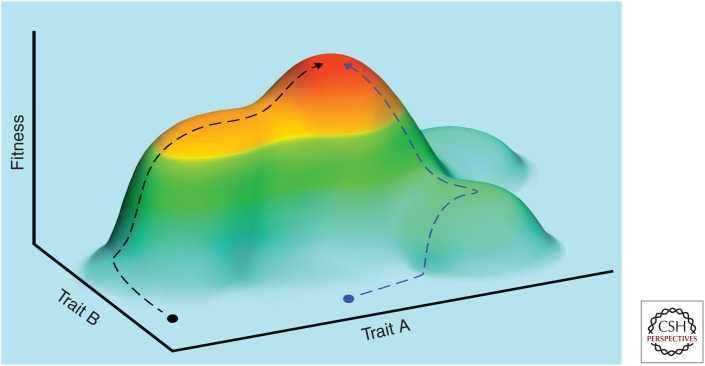
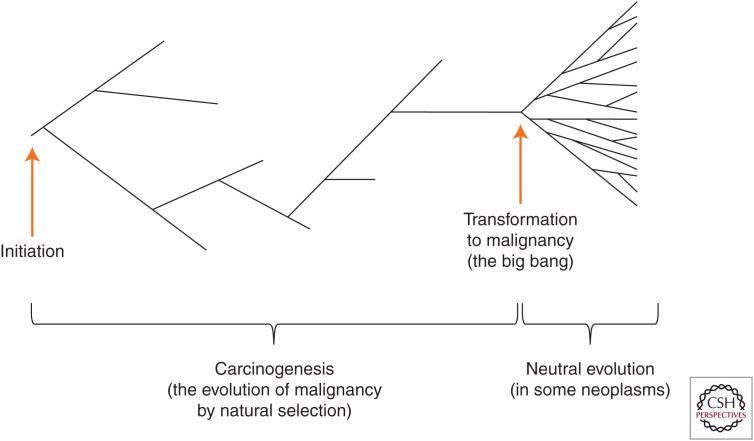
Evolution by natural selection is the conceptual foundation for nearly every branch of biology and increasingly also for biomedicine and medical research. In cancer biology, evolution explains how populations of cells in tumors change over time. It is a fundamental question whether this evolutionary process is driven primarily by natural selection and adaptation or by other evolutionary processes such as founder effects and drift. In cancer biology, as in organismal evolutionary biology, there is controversy about this question and also about the use of adaptation through natural selection as a guiding framework for research. In this review, we discuss the differences and similarities between evolution among somatic cells versus evolution among organisms. We review what is known about the parameters and rate of evolution in neoplasms, as well as evidence for adaptation. We conclude that adaptation is a useful framework that accurately explains the defining characteristics of cancer. Further, convergent evolution through natural selection provides the only satisfying explanation both for how a group of diverse pathologies have enough in common to usefully share the descriptive label of "cancer" and for why this convergent condition becomes life-threatening.
Copyright © 2017 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures

References
-
- An X, Tiwari AK, Sun Y, Ding PR, Ashby CR Jr, Chen ZS. 2010. BCR-ABL tyrosine kinase inhibitors in the treatment of Philadelphia chromosome positive chronic myeloid leukemia: A review. Leuk Res 34: 1255–1268. - PubMed
-
- Anderson K, Lutz C, van Delft FW, Bateman CM, Guo Y, Colman SM, Kempski H, Moorman AV, Titley I, Swansbury J, et al. 2011. Genetic variegation of clonal architecture and propagating cells in leukaemia. Nature 469: 356–361. - PubMed
-
- Araten DJ, Golde DW, Zhang RH, Thaler HT, Gargiulo L, Notaro R, Luzzatto L. 2005. A quantitative measurement of the human somatic mutation rate. Cancer Res 65: 8111–8117. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources