

Intein-mediated backbone cyclization of VP1 protein enhanced protection of CVB3-induced viral myocarditis
- PMID: 28148910
- PMCID: PMC5288654
- DOI: 10.1038/srep41485
Intein-mediated backbone cyclization of VP1 protein enhanced protection of CVB3-induced viral myocarditis
Abstract
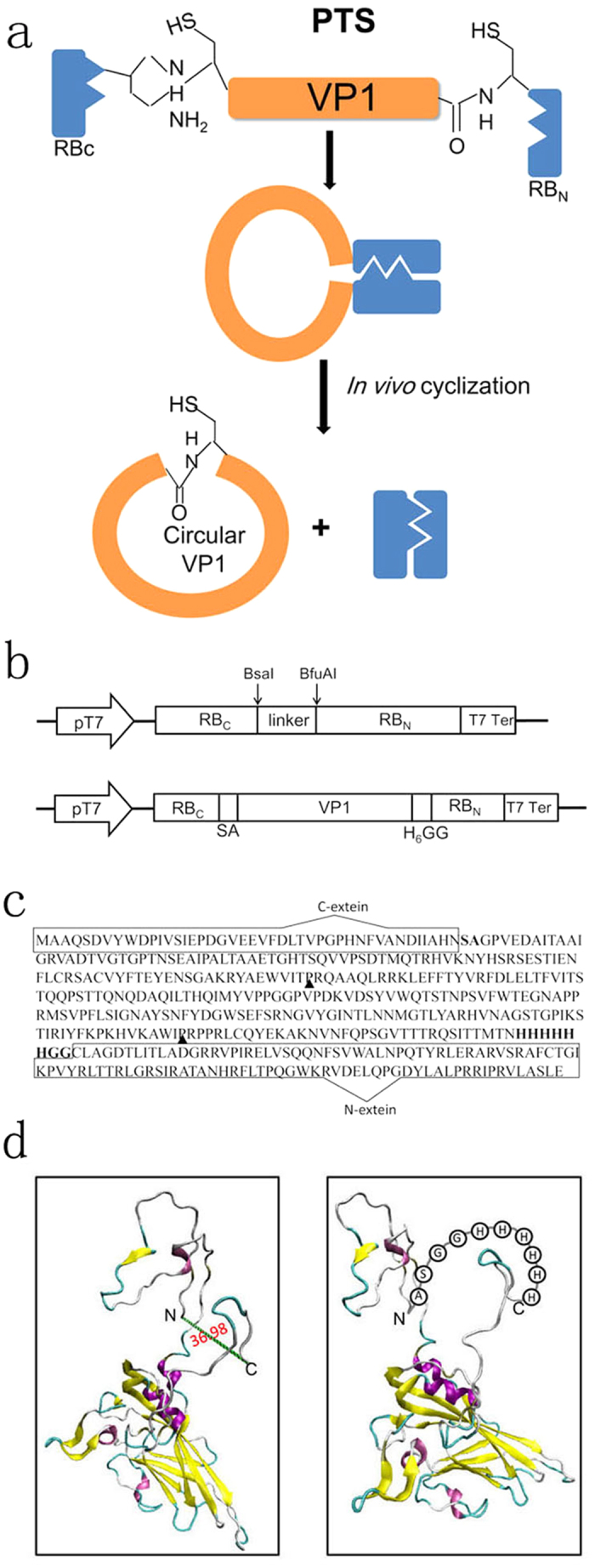
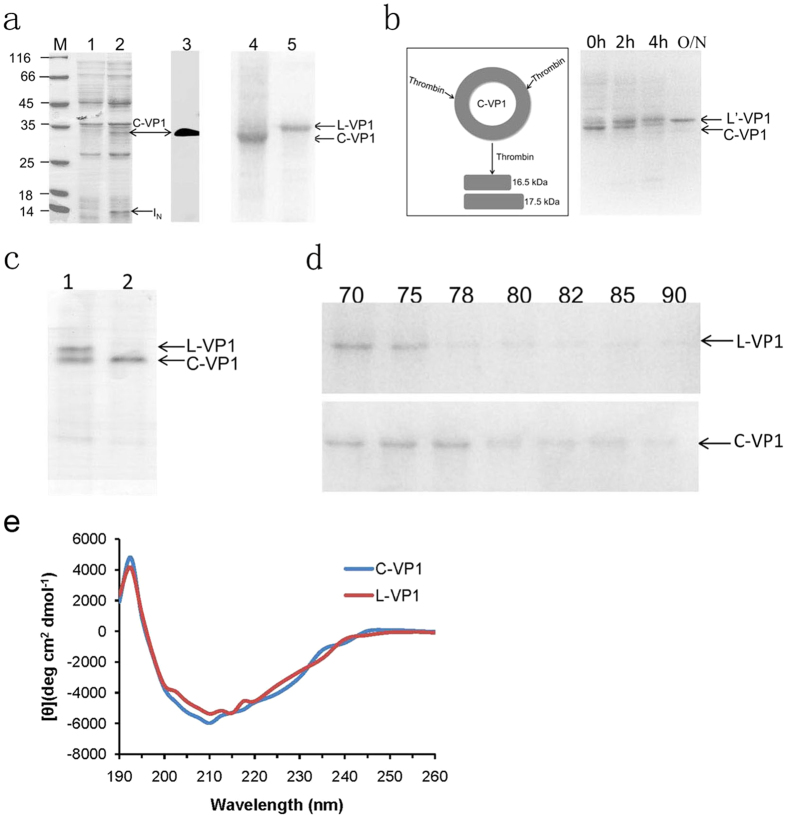
CVB3 is a common human pathogen to be highly lethal to newborns and causes viral myocarditis and pancreatitis in adults. However, there is no vaccine available for clinical use. CVB3 capsid protein VP1 is an immunodominant structural protein, containing several B- and T-cell epitopes. However, immunization of mice with VP1 protein is ineffective. Cyclization of peptide is commonly used to improve their in vivo stability and biological activity. Here, we designed and synthesizd cyclic VP1 protein by using engineered split Rma DnaB intein and the cyclization efficiency was 100% in E. coli. As a result, the cyclic VP1 was significantly more stable against irreversible aggregation upon heating and against carboxypeptidase in vitro and the degradation rate was more slowly in vivo. Compared with linear VP1, immunization mice with circular VP1 significantly increased CVB3-specific serum IgG level and augmented CVB3-specific cellular immune responses, consequently afforded better protection against CVB3-induced viral myocarditis. The cyclic VP1 may be a novel candidate protein vaccine for preventing CVB3 infection and similar approaches could be employed to a variety of protein vaccines to enhance their protection effect.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Spontaneous C-cleavage of a truncated intein as fusion tag to produce tag-free VP1 inclusion body nanoparticle vaccine against CVB3-induced viral myocarditis by the oral route.Microb Cell Fact. 2019 Apr 4;18(1):66. doi: 10.1186/s12934-019-1115-z. Microb Cell Fact. 2019. PMID: 30947747 Free PMC article.
-
AIM2 Co-immunization with VP1 Is Associated with Increased Memory CD8 T Cells and Mounts Long Lasting Protection against Coxsackievirus B3 Challenge.Front Cell Infect Microbiol. 2017 Jun 8;7:247. doi: 10.3389/fcimb.2017.00247. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 28642849 Free PMC article.
-
An Albumin-Binding Domain Peptide Confers Enhanced Immunoprotection Against Viral Myocarditis by CVB3 VP1 Vaccine.Front Immunol. 2021 Sep 22;12:666594. doi: 10.3389/fimmu.2021.666594. eCollection 2021. Front Immunol. 2021. PMID: 34630378 Free PMC article.
-
Manipulating intestinal immunity and microflora: an alternative solution to viral myocarditis?Future Microbiol. 2012 Oct;7(10):1207-16. doi: 10.2217/fmb.12.96. Future Microbiol. 2012. PMID: 23030425 Review.
-
Recombinant coxsackievirus vectors for prevention and therapy of virus-induced heart disease.Int J Med Microbiol. 2008 Jan;298(1-2):127-34. doi: 10.1016/j.ijmm.2007.08.010. Epub 2007 Sep 25. Int J Med Microbiol. 2008. PMID: 17897883 Review.
Cited by
-
Regulation of Apoptosis by Enteroviruses.Front Microbiol. 2020 Jun 3;11:1145. doi: 10.3389/fmicb.2020.01145. eCollection 2020. Front Microbiol. 2020. PMID: 32582091 Free PMC article. Review.
-
New Routes and Opportunities for Modular Construction of Particulate Vaccines: Stick, Click, and Glue.Front Immunol. 2018 Jun 26;9:1432. doi: 10.3389/fimmu.2018.01432. eCollection 2018. Front Immunol. 2018. PMID: 29997617 Free PMC article. Review.
-
Spontaneous C-cleavage of a truncated intein as fusion tag to produce tag-free VP1 inclusion body nanoparticle vaccine against CVB3-induced viral myocarditis by the oral route.Microb Cell Fact. 2019 Apr 4;18(1):66. doi: 10.1186/s12934-019-1115-z. Microb Cell Fact. 2019. PMID: 30947747 Free PMC article.
-
Covalently circularized nanodiscs; challenges and applications.Curr Opin Struct Biol. 2018 Aug;51:129-134. doi: 10.1016/j.sbi.2018.03.014. Epub 2018 Apr 20. Curr Opin Struct Biol. 2018. PMID: 29677570 Free PMC article. Review.
-
Intein-Mediated Protein Engineering for Cell-Based Biosensors.Biosensors (Basel). 2022 Apr 28;12(5):283. doi: 10.3390/bios12050283. Biosensors (Basel). 2022. PMID: 35624584 Free PMC article. Review.
References
-
- Maisch B., Ristic A. D., Hufnagel G. & Pankuweit S. Pathophysiology of viral myocarditis: the role of humoral immune response. Cardiovasc Pathol 11, 112–122 (2002). - PubMed
-
- Maisch B. D., Ristic A. D., Hufnagel G. & Pankuweit S. Pathophysiology of viral myocarditis - The role of humoral immune response. Cardiovascular Pathology 11, 112–122 (2002). - PubMed
-
- Kawai C. From myocarditis to cardiomyopathy: mechanisms of inflammation and cell death: learning from the past for the future. Circulation 99, 1091–1100 (1999). - PubMed
-
- Werner S., Klump W. M., Schonke H., Hofschneider P. H. & Kandolf R. Expression of coxsackievirus B3 capsid proteins in Escherichia coli and generation of virus-specific antisera. DNA 7, 307–316 (1988). - PubMed
-
- Huber S. et al.. T lymphocyte responses in CVB3-induced murine myocarditis. Scand J Infect Dis Suppl 88, 67–78 (1993). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
