

Robust network topologies for generating oscillations with temperature-independent periods
- PMID: 28152061
- PMCID: PMC5289577
- DOI: 10.1371/journal.pone.0171263
Robust network topologies for generating oscillations with temperature-independent periods
Abstract
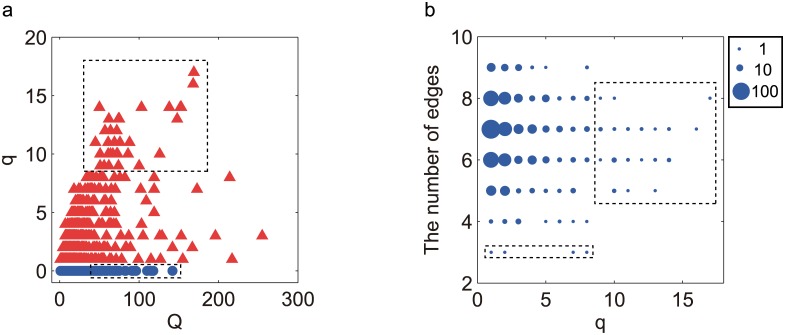
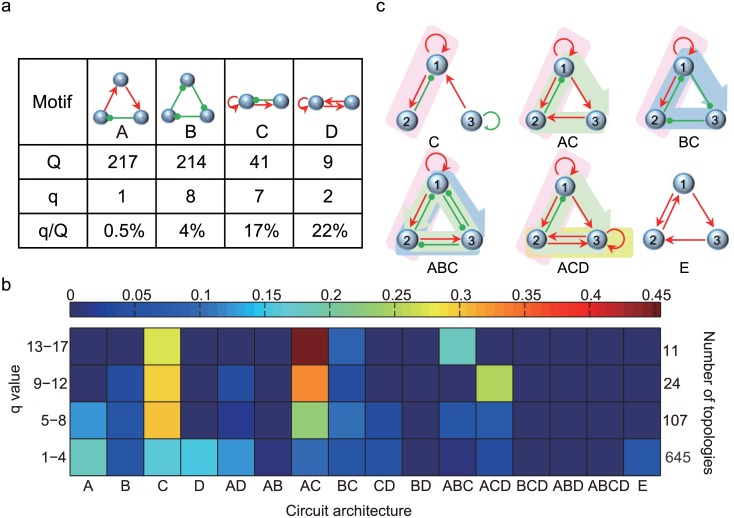
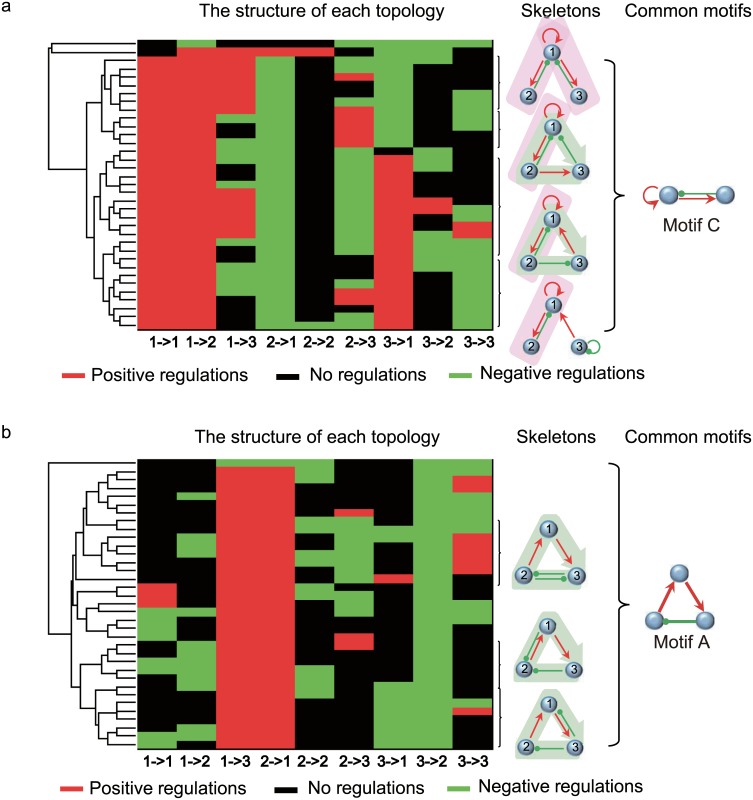
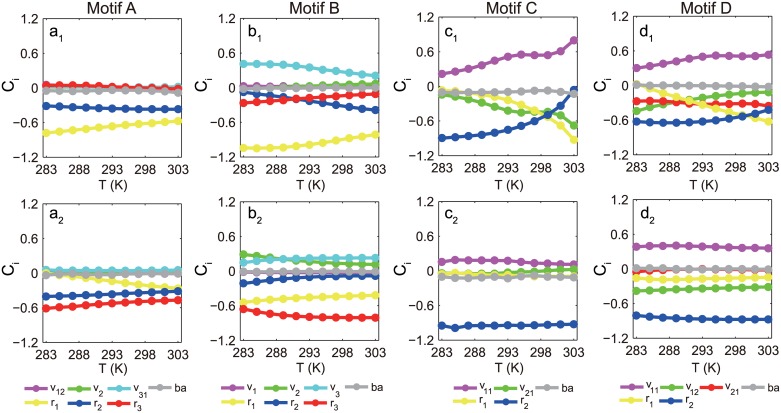
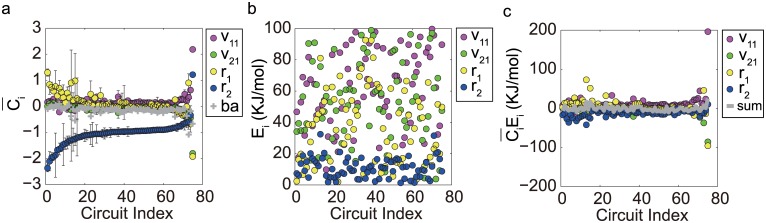
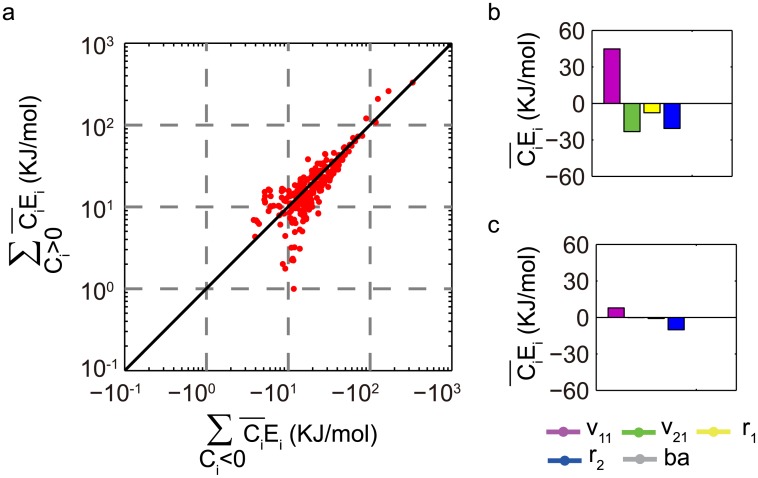
Nearly all living systems feature a temperature-independent oscillation period in circadian clocks. This ubiquitous property occurs at the system level and is rooted in the network architecture of the clock machinery. To investigate the mechanism of this prominent property of the circadian clock and provide general guidance for generating robust genetic oscillators with temperature-compensated oscillations, we theoretically explored the design principle and core network topologies preferred by oscillations with a temperature-independent period. By enumerating all topologies of genetic regulatory circuits with three genes, we obtained four network motifs, namely, a delayed negative feedback oscillator, repressilator, activator-inhibitor oscillator and substrate-depletion oscillator; hybrids of these motifs constitute the vast majority of target network topologies. These motifs are biased in their capacities for achieving oscillations and the temperature sensitivity of the period. The delayed negative feedback oscillator and repressilator are more robust for oscillations, whereas the activator-inhibitor and substrate-depletion oscillators are superior for maintaining a temperature-independent oscillation period. These results suggest that thermally robust oscillation can be more plausibly achieved by hybridizing these two categories of network motifs. Antagonistic balance and temperature insulation mechanisms for achieving temperature compensation are typically found in these topologies with temperature robustness. In the temperature insulation approach, the oscillation period relies on very few parameters, and these parameters are influenced only slightly by temperature. This approach prevents the temperature from affecting the oscillation period and generates circadian rhythms that are robust against environmental perturbations.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Analytical study of robustness of a negative feedback oscillator by multiparameter sensitivity.BMC Syst Biol. 2014;8 Suppl 5(Suppl 5):S1. doi: 10.1186/1752-0509-8-S5-S1. Epub 2014 Dec 12. BMC Syst Biol. 2014. PMID: 25605374 Free PMC article.
-
Systematic analysis of negative and positive feedback loops for robustness and temperature compensation in circadian rhythms.NPJ Syst Biol Appl. 2023 Feb 11;9(1):5. doi: 10.1038/s41540-023-00268-7. NPJ Syst Biol Appl. 2023. PMID: 36774353 Free PMC article.
-
A design principle for a posttranslational biochemical oscillator.Cell Rep. 2012 Oct 25;2(4):938-50. doi: 10.1016/j.celrep.2012.09.006. Epub 2012 Oct 19. Cell Rep. 2012. PMID: 23084745
-
The design of intracellular oscillators that interact with metabolism.Cell Mol Life Sci. 2006 Jun;63(11):1215-20. doi: 10.1007/s00018-005-5611-4. Cell Mol Life Sci. 2006. PMID: 16649146 Free PMC article. Review.
-
Biological timing and the clock metaphor: oscillatory and hourglass mechanisms.Chronobiol Int. 2001 May;18(3):329-69. doi: 10.1081/cbi-100103961. Chronobiol Int. 2001. PMID: 11475408 Review.
Cited by
-
Cut the noise or couple up: Coordinating circadian and synthetic clocks.iScience. 2021 Aug 27;24(9):103051. doi: 10.1016/j.isci.2021.103051. eCollection 2021 Sep 24. iScience. 2021. PMID: 34568785 Free PMC article. Review.
-
The Topological Characteristics of Biological Ratio-Sensing Networks.Life (Basel). 2023 Jan 28;13(2):351. doi: 10.3390/life13020351. Life (Basel). 2023. PMID: 36836707 Free PMC article.
-
Temperature compensation through kinetic regulation in biochemical oscillators.Proc Natl Acad Sci U S A. 2024 May 21;121(21):e2401567121. doi: 10.1073/pnas.2401567121. Epub 2024 May 15. Proc Natl Acad Sci U S A. 2024. PMID: 38748573 Free PMC article.
-
Robust network topologies for temperature-inducible bioswitches.J Biol Eng. 2022 May 23;16(1):12. doi: 10.1186/s13036-022-00290-z. J Biol Eng. 2022. PMID: 35606858 Free PMC article.
-
Molecular Mechanisms Regulating Temperature Compensation of the Circadian Clock.Front Neurol. 2017 Apr 27;8:161. doi: 10.3389/fneur.2017.00161. eCollection 2017. Front Neurol. 2017. PMID: 28496429 Free PMC article. Review.
References
-
- Denlinger DL, Giebultowicz J, Saunders DS. Insect timing: circadian rhythmicity to seasonality: Elsevier; 2001.
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
