

Coupling organelle inheritance with mitosis to balance growth and differentiation
- PMID: 28154022
- PMCID: PMC5333555
- DOI: 10.1126/science.aah4701
Coupling organelle inheritance with mitosis to balance growth and differentiation
Abstract
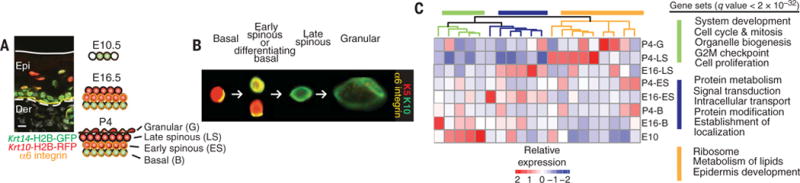
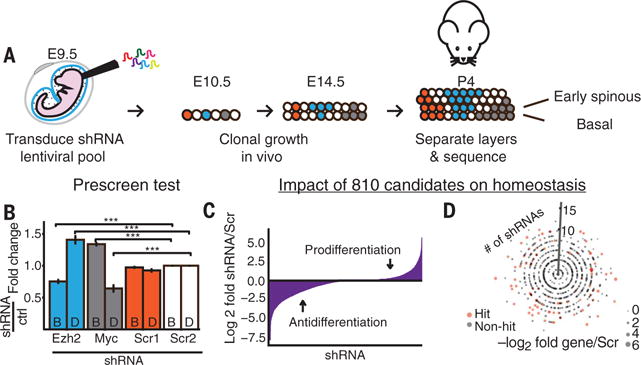
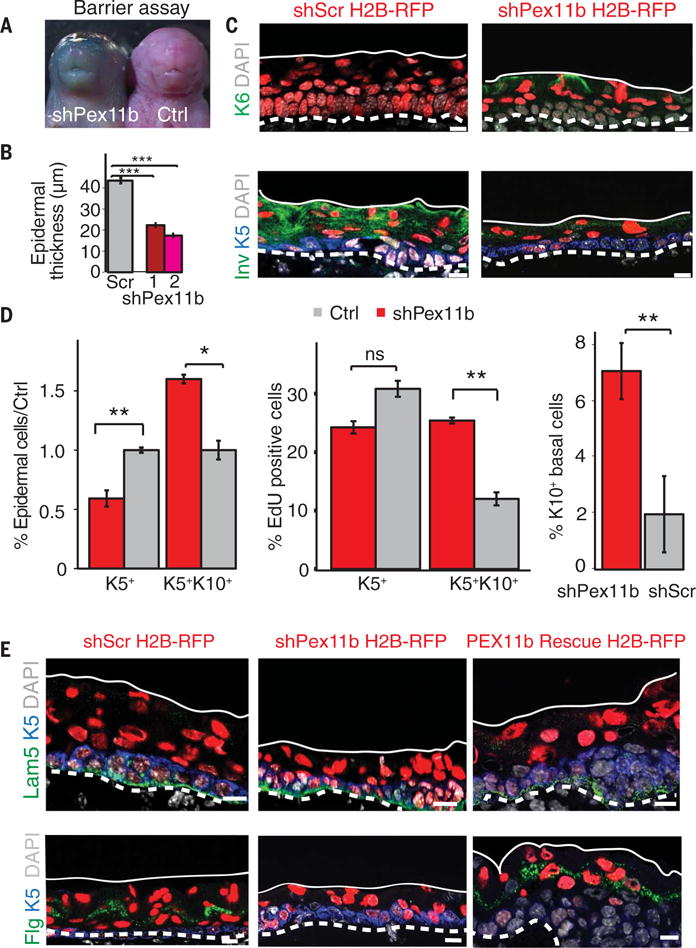
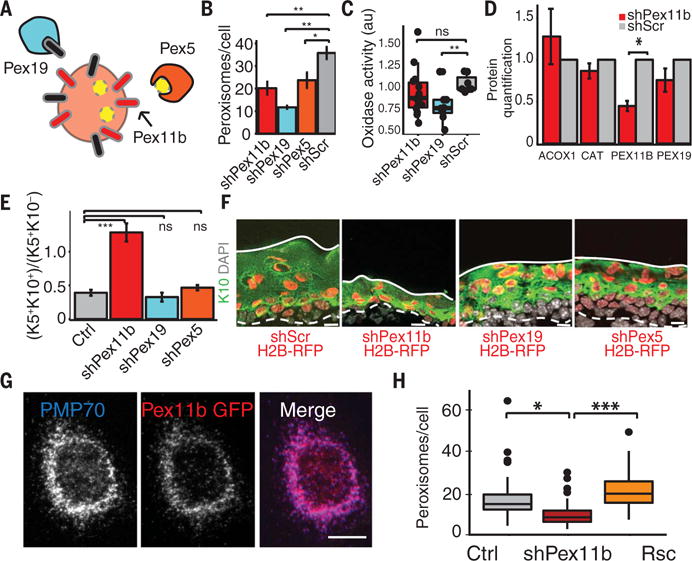
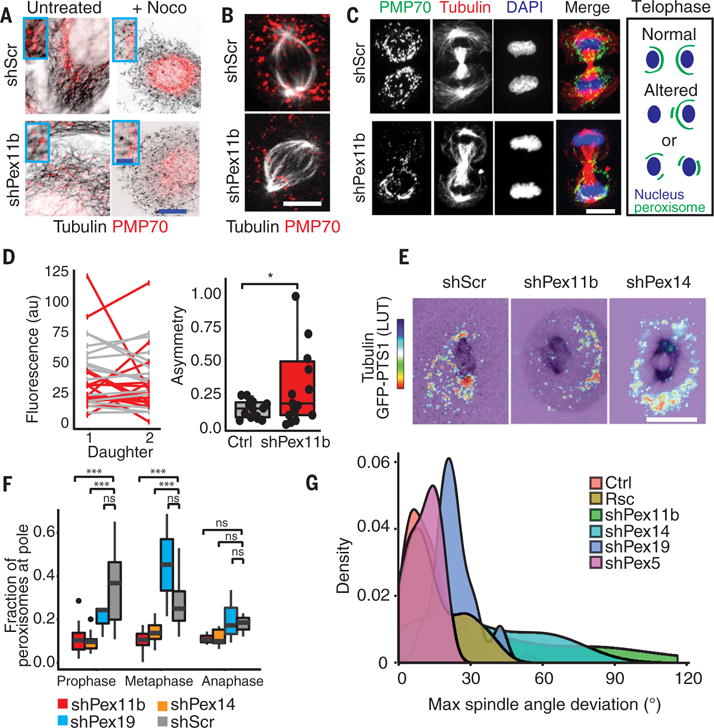
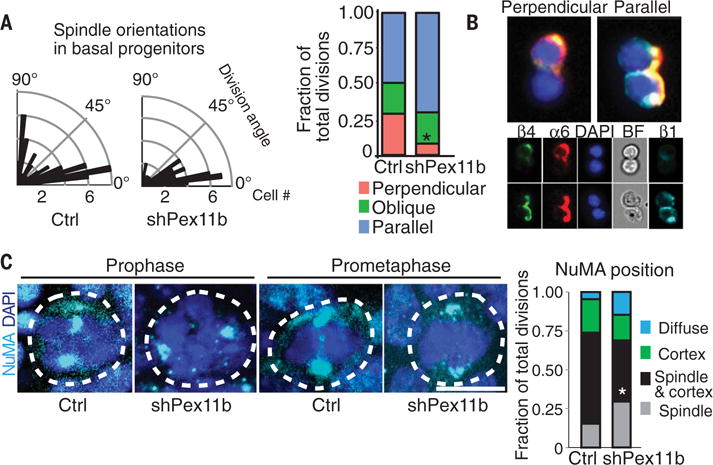
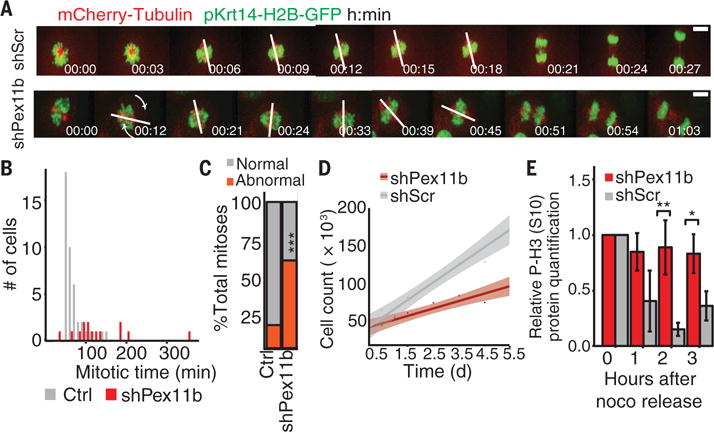
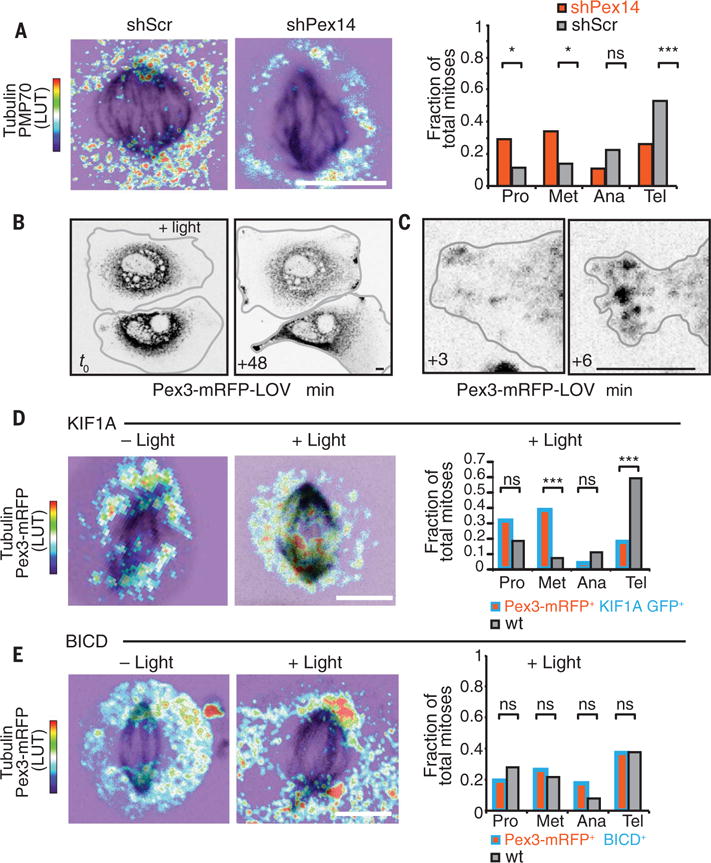
Balancing growth and differentiation is essential to tissue morphogenesis and homeostasis. How imbalances arise in disease states is poorly understood. To address this issue, we identified transcripts differentially expressed in mouse basal epidermal progenitors versus their differentiating progeny and those altered in cancers. We used an in vivo RNA interference screen to unveil candidates that altered the equilibrium between the basal proliferative layer and suprabasal differentiating layers forming the skin barrier. We found that epidermal progenitors deficient in the peroxisome-associated protein Pex11b failed to segregate peroxisomes properly and entered a mitotic delay that perturbed polarized divisions and skewed daughter fates. Together, our findings unveil a role for organelle inheritance in mitosis, spindle alignment, and the choice of daughter progenitors to differentiate or remain stem-like.
Copyright © 2017, American Association for the Advancement of Science.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
Organelle inheritance-what players have skin in the game?Science. 2017 Feb 3;355(6324):459-460. doi: 10.1126/science.aam5981. Science. 2017. PMID: 28154036 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
