

Ultrastructural evidence for synaptic scaling across the wake/sleep cycle
- PMID: 28154076
- PMCID: PMC5313037
- DOI: 10.1126/science.aah5982
Ultrastructural evidence for synaptic scaling across the wake/sleep cycle
Abstract
It is assumed that synaptic strengthening and weakening balance throughout learning to avoid runaway potentiation and memory interference. However, energetic and informational considerations suggest that potentiation should occur primarily during wake, when animals learn, and depression should occur during sleep. We measured 6920 synapses in mouse motor and sensory cortices using three-dimensional electron microscopy. The axon-spine interface (ASI) decreased ~18% after sleep compared with wake. This decrease was proportional to ASI size, which is indicative of scaling. Scaling was selective, sparing synapses that were large and lacked recycling endosomes. Similar scaling occurred for spine head volume, suggesting a distinction between weaker, more plastic synapses (~80%) and stronger, more stable synapses. These results support the hypothesis that a core function of sleep is to renormalize overall synaptic strength increased by wake.
Copyright © 2017, American Association for the Advancement of Science.
Figures
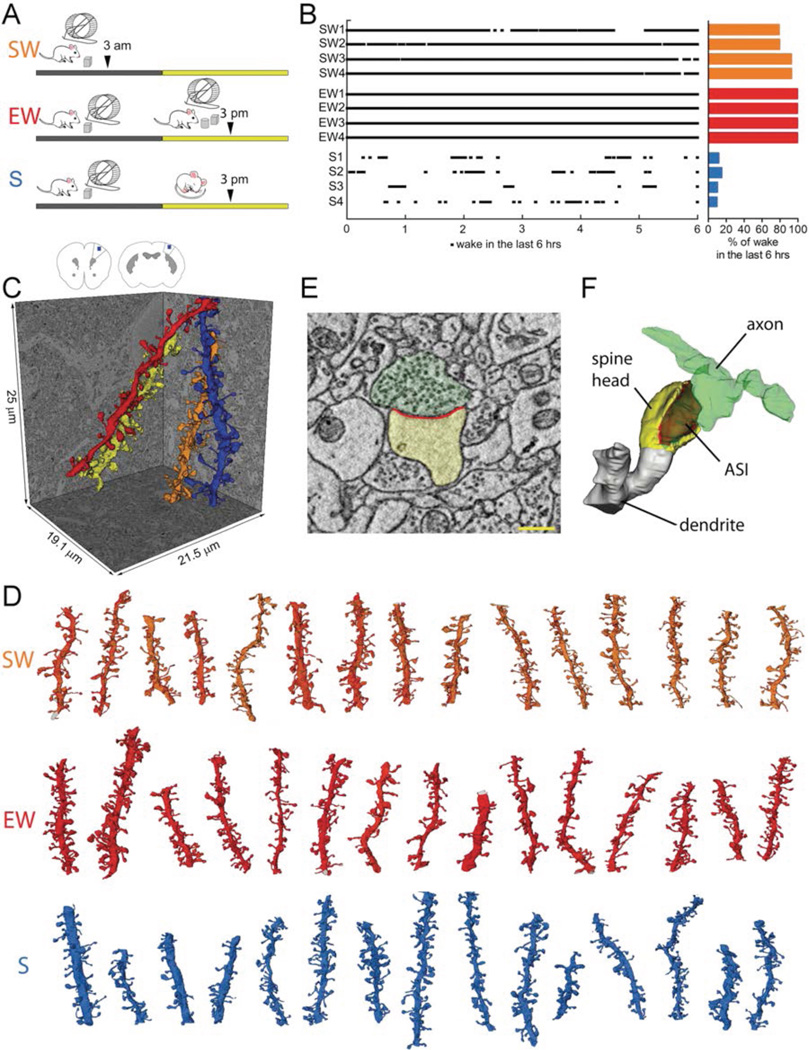
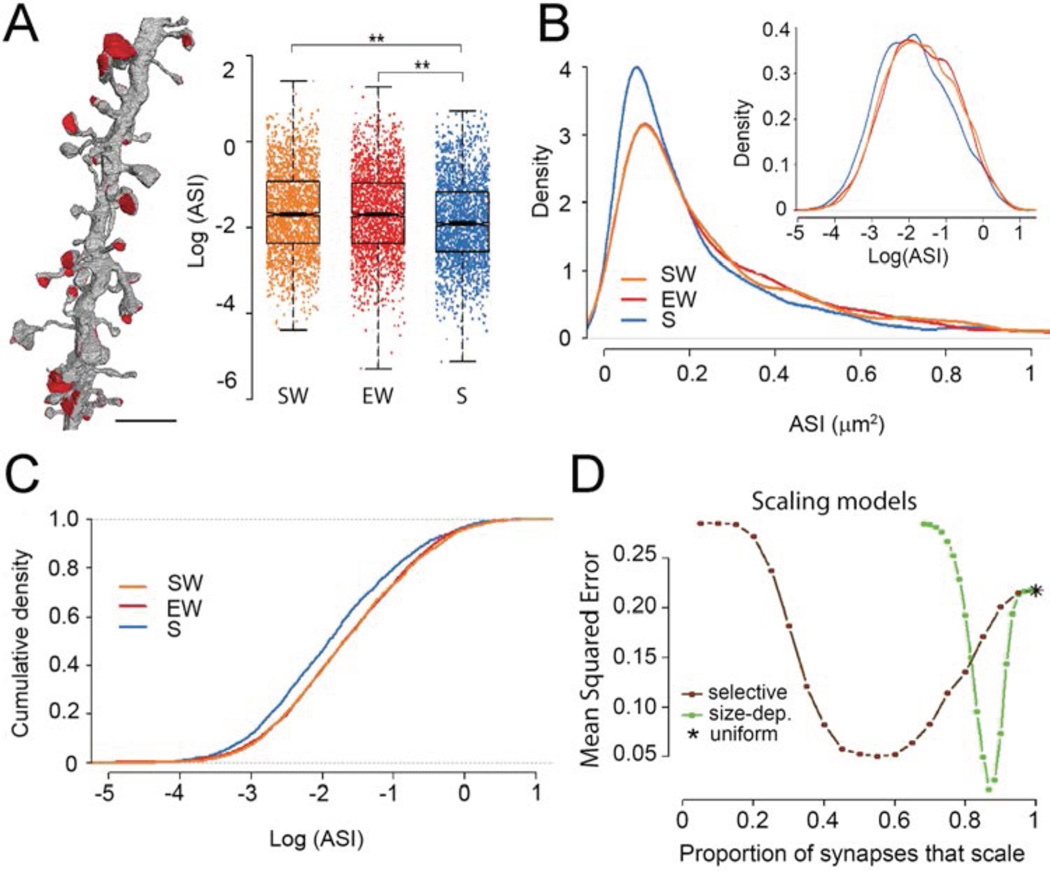
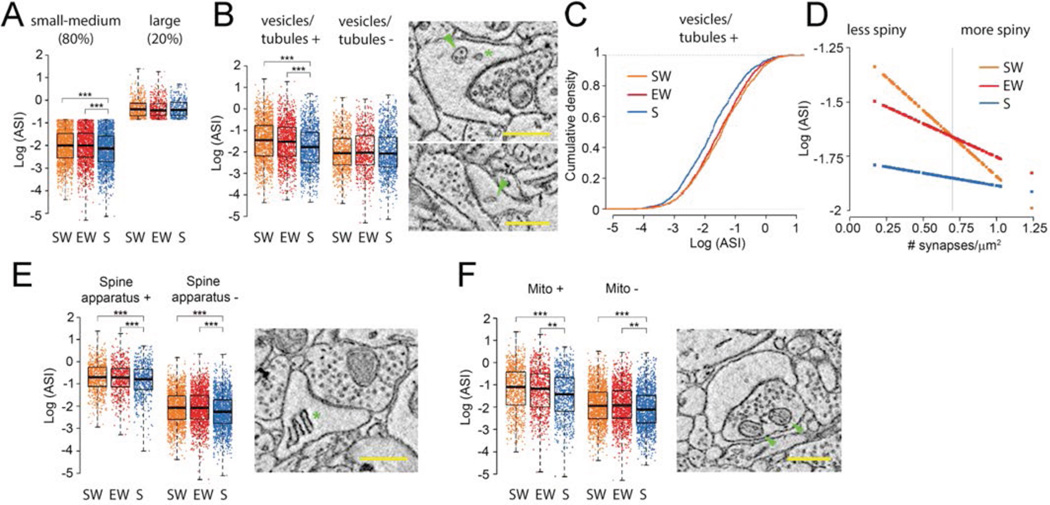
Comment in
-
Homeostasis of Synapses: Expansion During Wakefulness, Contraction During Sleep.Neurosci Bull. 2017 Jun;33(3):359-360. doi: 10.1007/s12264-017-0130-5. Epub 2017 Apr 7. Neurosci Bull. 2017. PMID: 28389869 Free PMC article. No abstract available.
Dataset use reported in
-
Synaptic scaling in sleep.Science. 2017 Feb 3;355(6324):457. doi: 10.1126/science.aam7917. Science. 2017. PMID: 28154034 No abstract available.
References
-
- Tang Y, Nyengaard JR, De Groot DM, Gundersen HJ. Total regional and global number of synapses in the human brain neocortex. Synapse. 2001;41:258–273. - PubMed
-
- Holtmaat A, Svoboda K. Experience-dependent structural synaptic plasticity in the mammalian brain. Nat Rev Neurosci. 2009;10:647–658. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
