

Not All H3K4 Methylations Are Created Equal: Mll2/COMPASS Dependency in Primordial Germ Cell Specification
- PMID: 28157506
- PMCID: PMC5404503
- DOI: 10.1016/j.molcel.2017.01.013
Not All H3K4 Methylations Are Created Equal: Mll2/COMPASS Dependency in Primordial Germ Cell Specification
Abstract
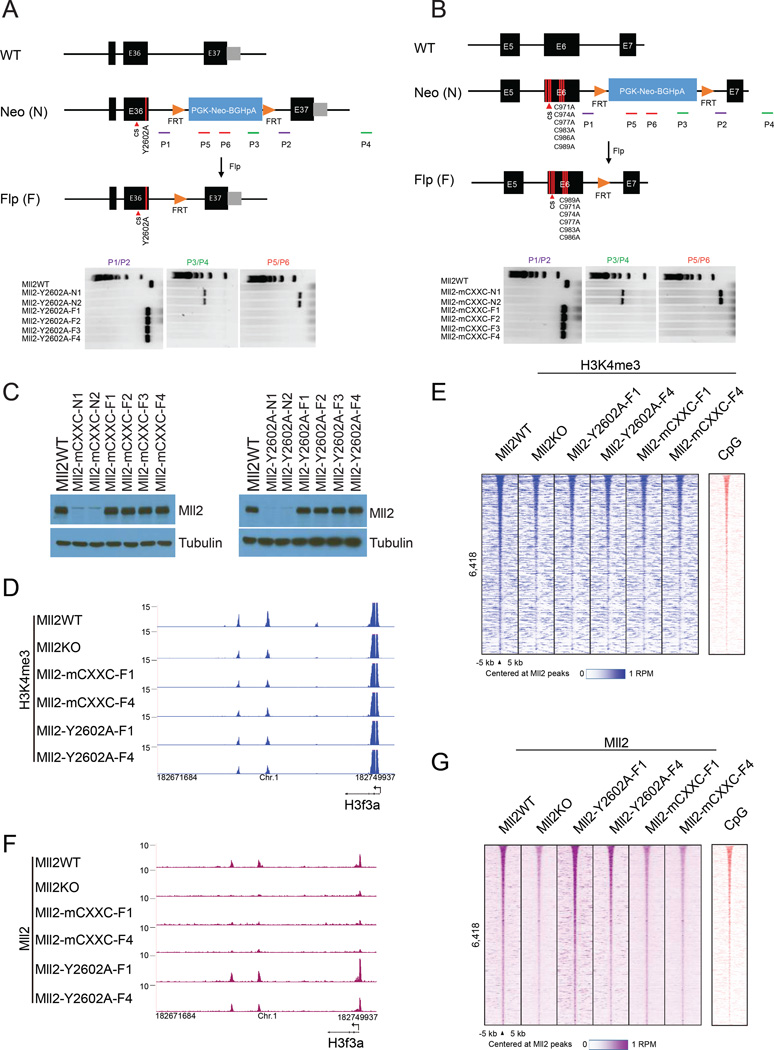
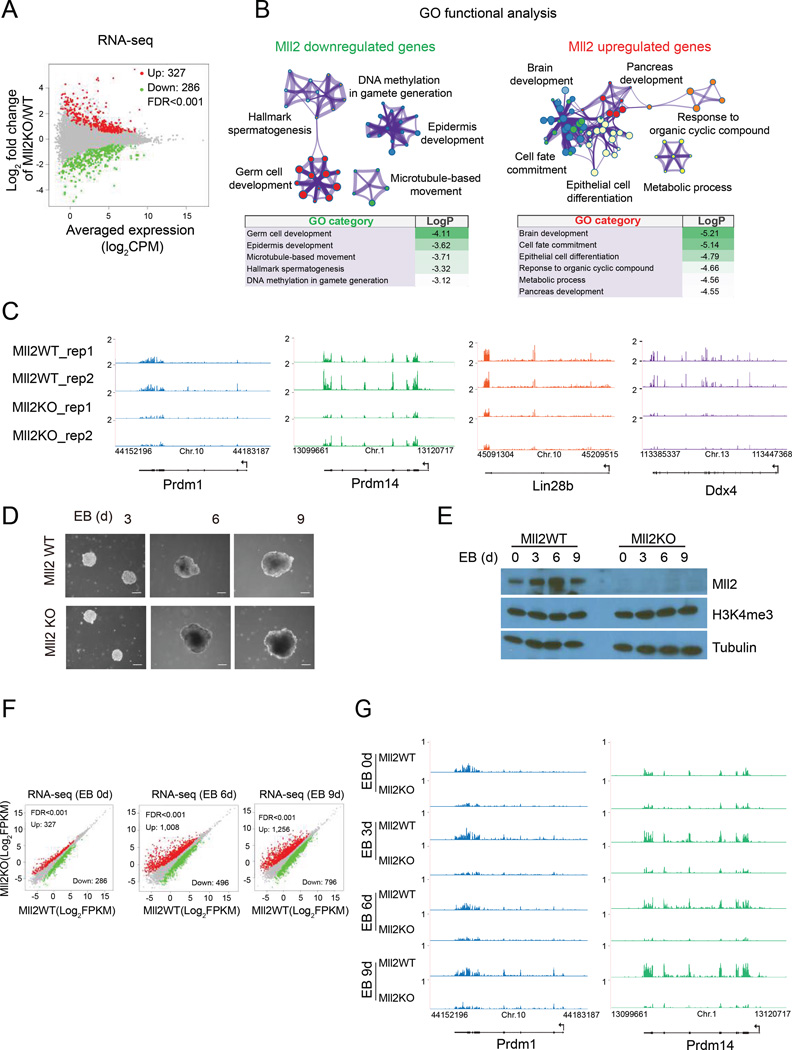
The spatiotemporal regulation of gene expression is central for cell-lineage specification during embryonic development and is achieved through the combinatorial action of transcription factors/co-factors and epigenetic states at cis-regulatory elements. Here, we show that in addition to implementing H3K4me3 at promoters of bivalent genes, Mll2 (KMT2B)/COMPASS can also implement H3K4me3 at a subset of non-TSS regulatory elements, a subset of which shares epigenetic signatures of active enhancers. Our mechanistic studies reveal that association of Mll2's CXXC domain with CpG-rich regions plays an instrumental role for chromatin targeting and subsequent implementation of H3K4me3. Although Mll2/COMPASS is required for H3K4me3 implementation on thousands of loci, generation of catalytically mutant MLL2/COMPASS demonstrated that H3K4me3 implemented by this enzyme was essential for expression of a subset of genes, including those functioning in the control of transcriptional programs during embryonic development. Our findings suggest that not all H3K4 trimethylations implemented by MLL2/COMPASS are functionally equivalent.
Keywords: CXXC domain; Histone-Lysine N-Methyltransferase; Kmt2b; Mll2; chromatin; embryonic development; gene expression regulation; histone; mouse embryonic stem cell; primordial germ cell.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures





References
-
- Azuara V, Perry P, Sauer S, Spivakov M, Jorgensen HF, John RM, Gouti M, Casanova M, Warnes G, Merkenschlager M, et al. Chromatin signatures of pluripotent cell lines. Nat Cell Biol. 2006;8:532–538. - PubMed
-
- Bernstein BE, Mikkelsen TS, Xie X, Kamal M, Huebert DJ, Cuff J, Fry B, Meissner A, Wernig M, Plath K, et al. A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell. 2006;125:315–326. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
