

The lncRNA VELUCT strongly regulates viability of lung cancer cells despite its extremely low abundance
- PMID: 28160600
- PMCID: PMC5435915
- DOI: 10.1093/nar/gkx076
The lncRNA VELUCT strongly regulates viability of lung cancer cells despite its extremely low abundance
Abstract
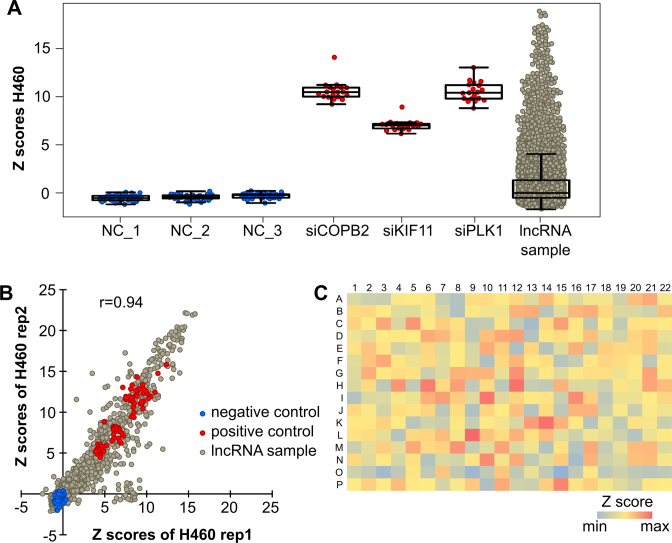
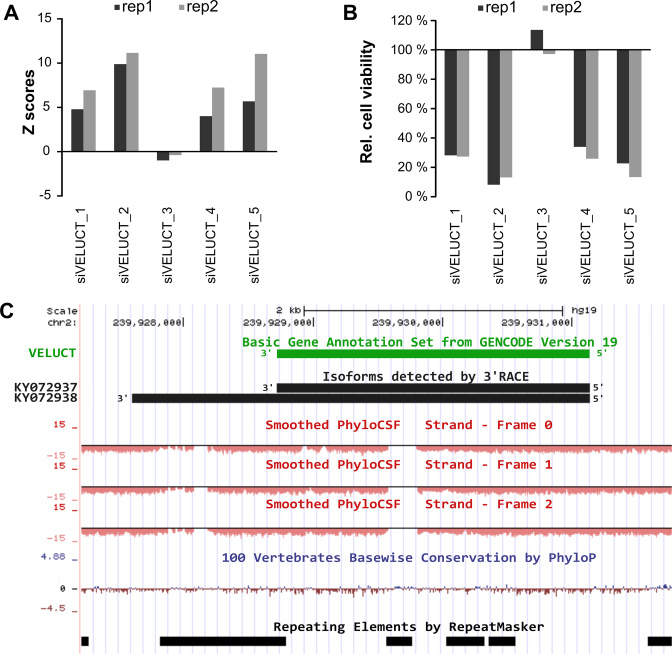
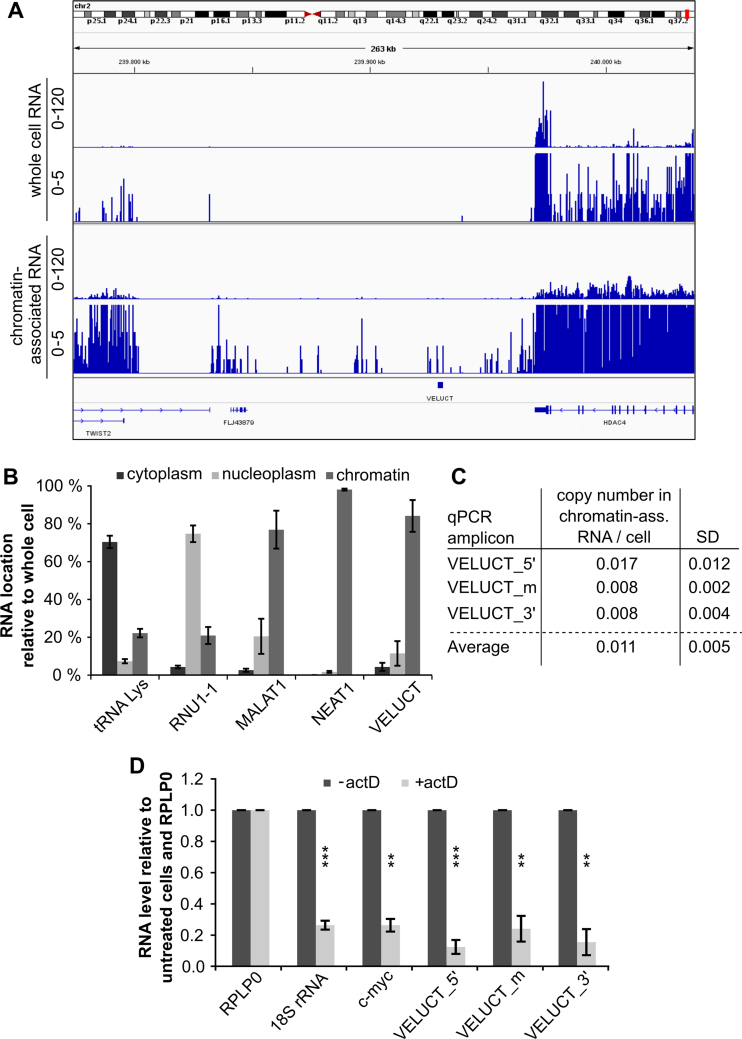
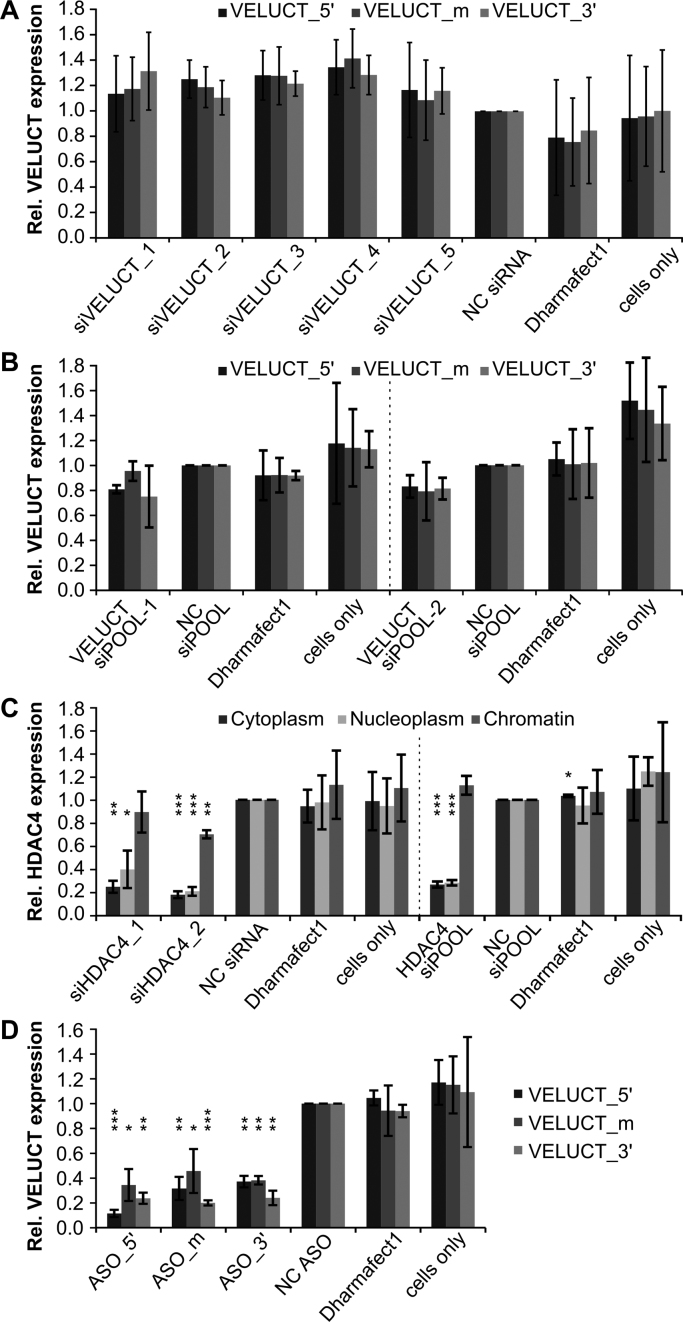
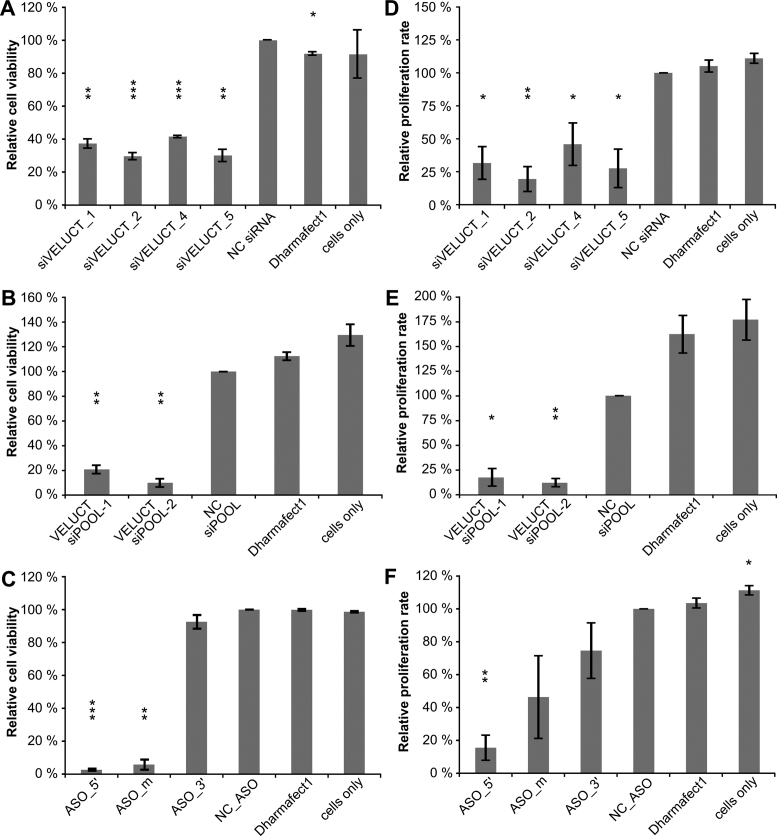
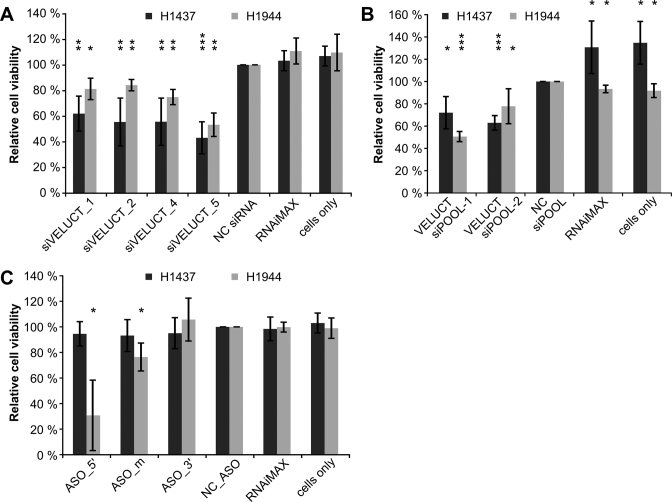
Little is known about the function of most non-coding RNAs (ncRNAs). The majority of long ncRNAs (lncRNAs) is expressed at very low levels and it is a matter of intense debate whether these can be of functional relevance. Here, we identified lncRNAs regulating the viability of lung cancer cells in a high-throughput RNA interference screen. Based on our previous expression profiling, we designed an siRNA library targeting 638 lncRNAs upregulated in human cancer. In a functional siRNA screen analyzing the viability of lung cancer cells, the most prominent hit was a novel lncRNA which we called Viability Enhancing LUng Cancer Transcript (VELUCT). In silico analyses confirmed the non-coding properties of the transcript. Surprisingly, VELUCT was below the detection limit in total RNA from NCI-H460 cells by RT-qPCR as well as RNA-Seq, but was robustly detected in the chromatin-associated RNA fraction. It is an extremely low abundant lncRNA with an RNA copy number of less than one copy per cell. Blocking transcription with actinomycin D revealed that VELUCT RNA was highly unstable which may partially explain its low steady-state concentration. Despite its extremely low abundance, loss-of-function of VELUCT with three independent experimental approaches in three different lung cancer cell lines led to a significant reduction of cell viability: Next to four individual siRNAs, also two complex siPOOLs as well as two antisense oligonucleotides confirmed the strong and specific phenotype. In summary, the extremely low abundant lncRNA VELUCT is essential for regulation of cell viability in several lung cancer cell lines. Hence, VELUCT is the first example for a lncRNA that is expressed at a very low level, but has a strong loss-of-function phenotype. Thus, our study proves that at least individual low-abundant lncRNAs can play an important functional role.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Comment in
-
VELUCT, a long non-coding RNA with an important cellular function despite low abundance.J Thorac Dis. 2017 Oct;9(10):3638-3640. doi: 10.21037/jtd.2017.09.66. J Thorac Dis. 2017. PMID: 29268362 Free PMC article. No abstract available.
References
-
- Kapranov P., Cheng J., Dike S., Nix D.A., Duttagupta R., Willingham A.T., Stadler P.F., Hertel J., Hackermuller J., Hofacker I.L. et al. RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science. 2007; 316:1484–1488. - PubMed
-
- Yan L., Yang M., Guo H., Yang L., Wu J., Li R., Liu P., Lian Y., Zheng X., Yan J. et al. Single-cell RNA-Seq profiling of human preimplantation embryos and embryonic stem cells. Nat. Struct. Mol. Biol. 2013; 20:1131–1139. - PubMed
-
- Tsoi L.C., Iyer M.K., Stuart P.E., Swindell W.R., Gudjonsson J.E., Tejasvi T., Sarkar M.K., Li B., Ding J., Voorhees J.J. et al. Analysis of long non-coding RNAs highlights tissue-specific expression patterns and epigenetic profiles in normal and psoriatic skin. Genome Biol. 2015; 16:24. - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
