

Axonal Endoplasmic Reticulum Ca2+ Content Controls Release Probability in CNS Nerve Terminals
- PMID: 28162809
- PMCID: PMC5325711
- DOI: 10.1016/j.neuron.2017.01.010
Axonal Endoplasmic Reticulum Ca2+ Content Controls Release Probability in CNS Nerve Terminals
Abstract
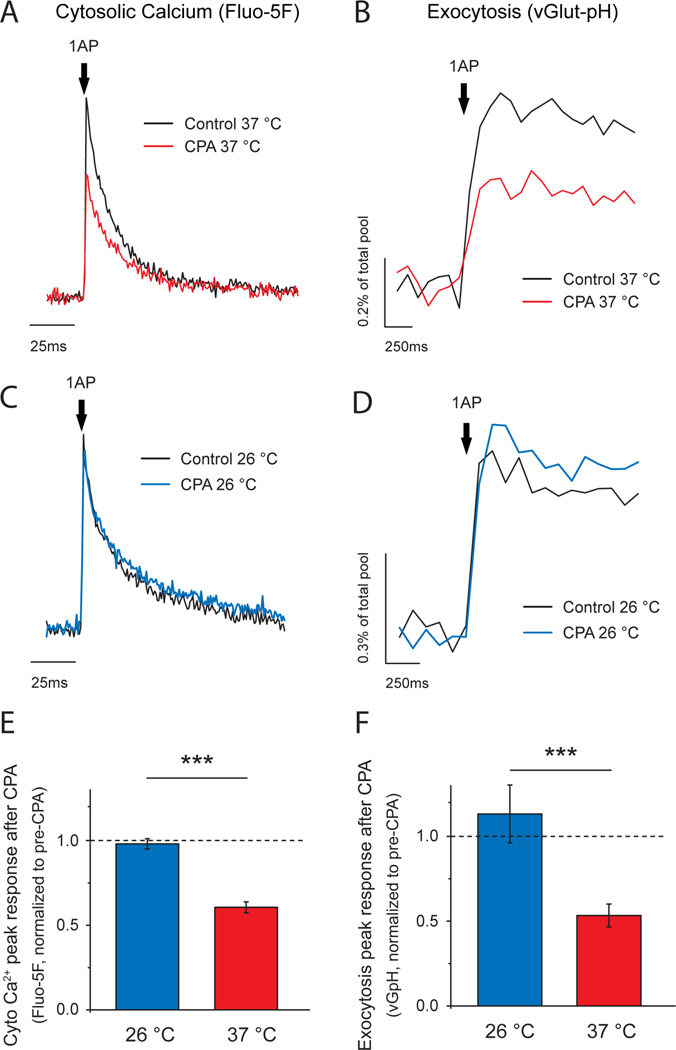
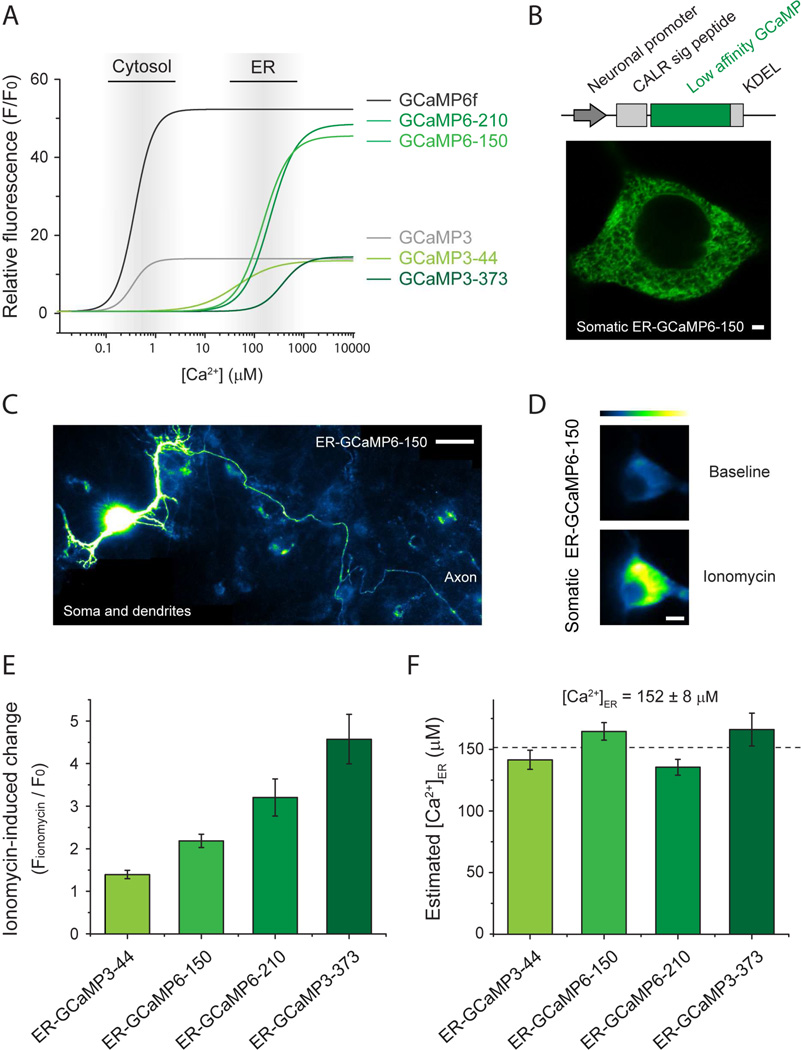
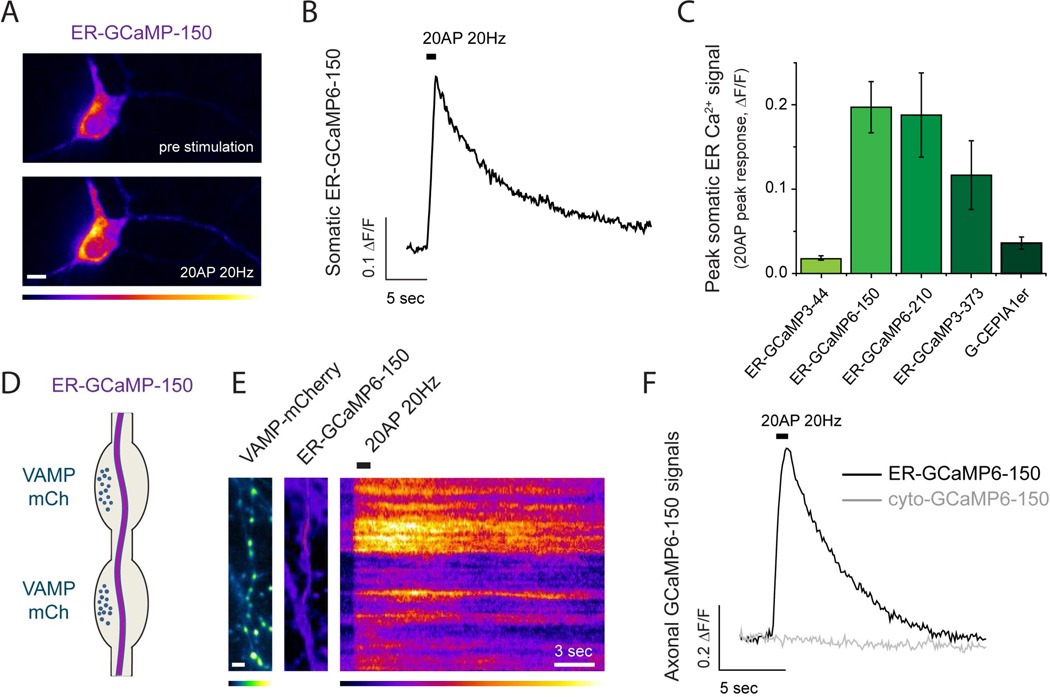
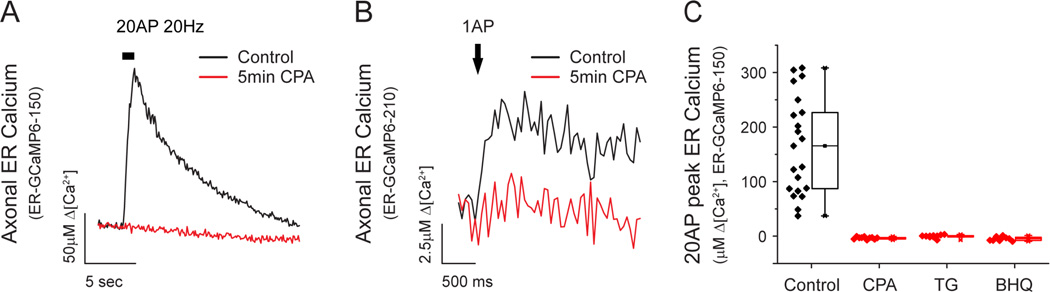
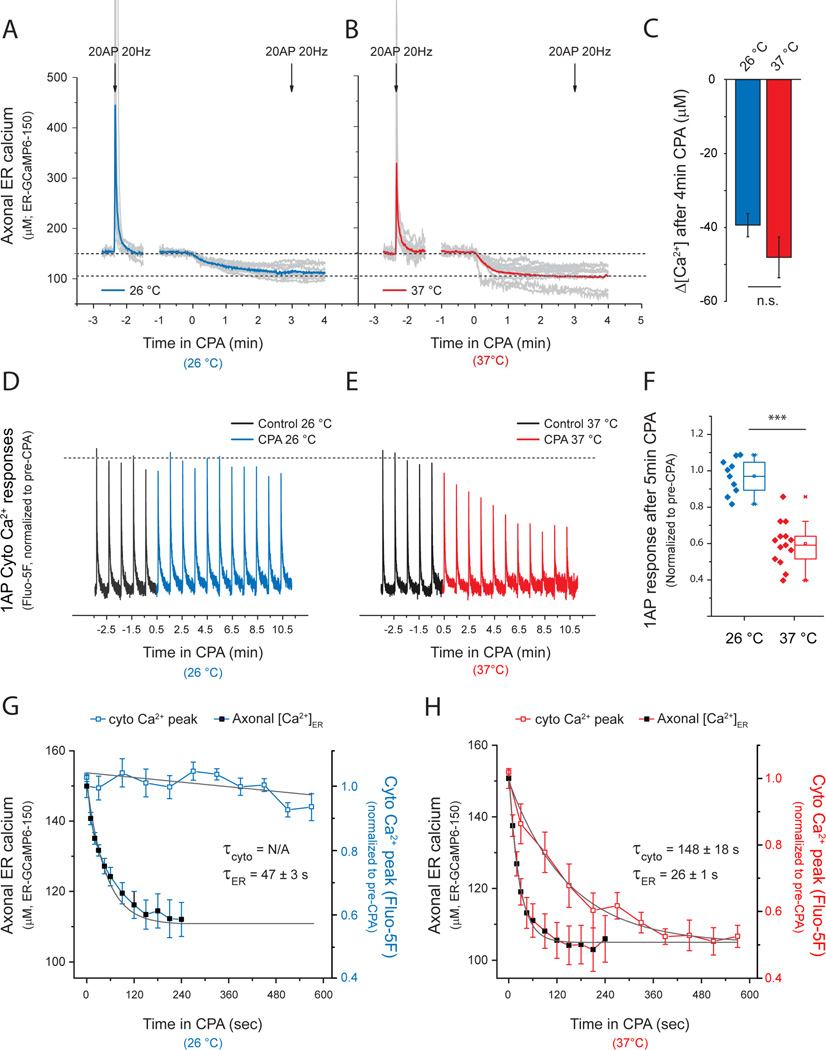
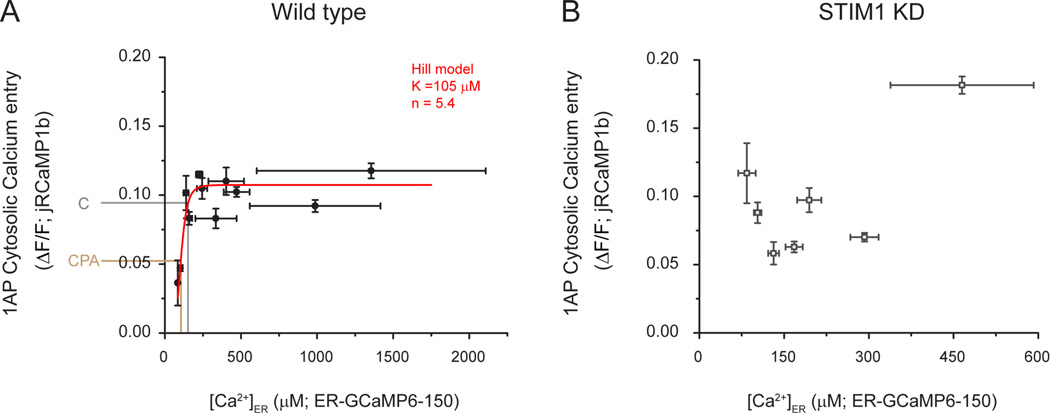
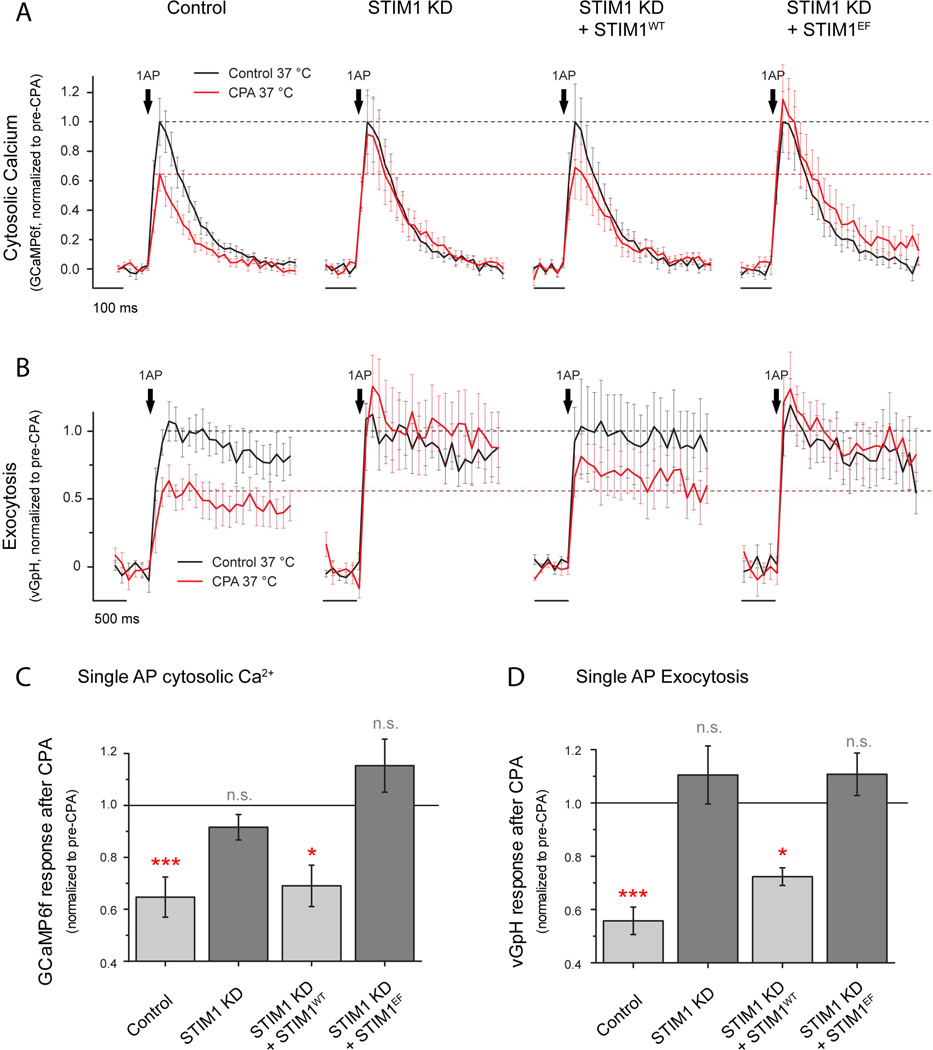
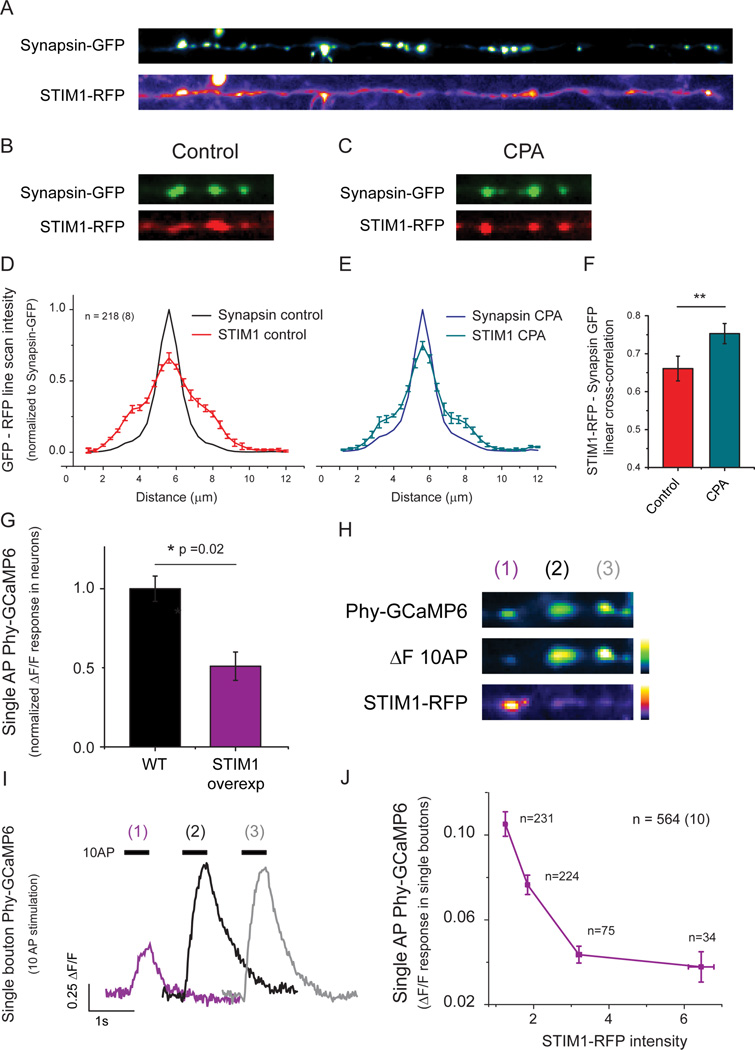
Although the endoplasmic reticulum (ER) extends throughout axons and axonal ER dysfunction is implicated in numerous neurological diseases, its role at nerve terminals is poorly understood. We developed novel genetically encoded ER-targeted low-affinity Ca2+ indicators optimized for examining axonal ER Ca2+. Our experiments revealed that presynaptic function is tightly controlled by ER Ca2+ content. We found that neuronal activity drives net Ca2+ uptake into presynaptic ER although this activity does not contribute significantly to shaping cytosolic Ca2+ except during prolonged repetitive firing. In contrast, we found that axonal ER acts as an actuator of plasma membrane (PM) function: [Ca2+]ER controls STIM1 activation in presynaptic terminals, which results in the local modulation of presynaptic function, impacting activity-driven Ca2+ entry and release probability. These experiments reveal a critical role of presynaptic ER in the control of neurotransmitter release and will help frame future investigations into the molecular basis of ER-driven neuronal disease states.
Keywords: Ca(2+) imaging; ER-GCaMP; GCaMP6; STIM1; low affinity Ca(2+) indicator; presynaptic endoplasmic reticulum.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Alonso MT, Barrero MJ, Carnicero E, Montero M, Garcia-Sancho J, Alvarez J. Functional measurements of [Ca2+] in the endoplasmic reticulum using a herpes virus to deliver targeted aequorin. Cell Calcium. 1998;24:87–96. - PubMed
-
- Beck A, Lohr C, Deitmer JW. Calcium transients in subcompartments of the leech Retzius neuron as induced by single action potentials. J Neurobiol. 2001;48:1–18. - PubMed
-
- Berna-Erro A, Braun A, Kraft R, Kleinschnitz C, Schuhmann MK, Stegner D, Wultsch T, Eilers J, Meuth SG, Stoll G, et al. STIM2 regulates capacitive Ca2+ entry in neurons and plays a key role in hypoxic neuronal cell death. Sci Signal. 2009;2:ra67. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
