

Parathyroid Hormone Directs Bone Marrow Mesenchymal Cell Fate
- PMID: 28162969
- PMCID: PMC5342925
- DOI: 10.1016/j.cmet.2017.01.001
Parathyroid Hormone Directs Bone Marrow Mesenchymal Cell Fate
Abstract
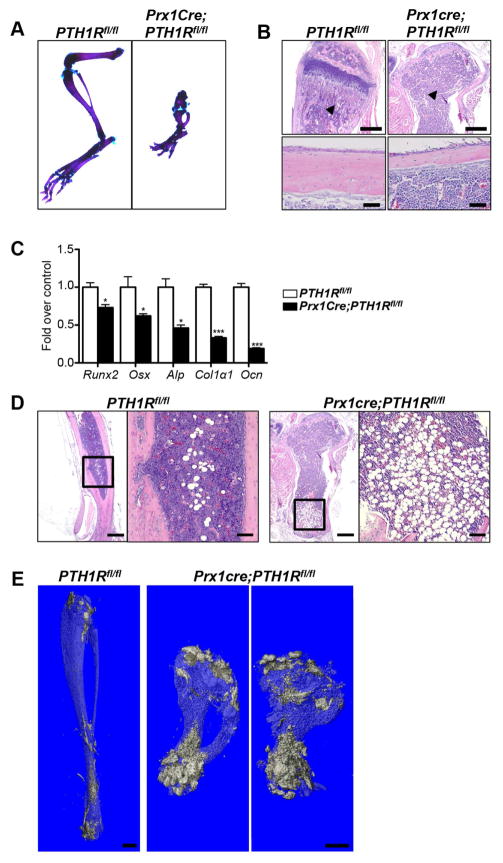
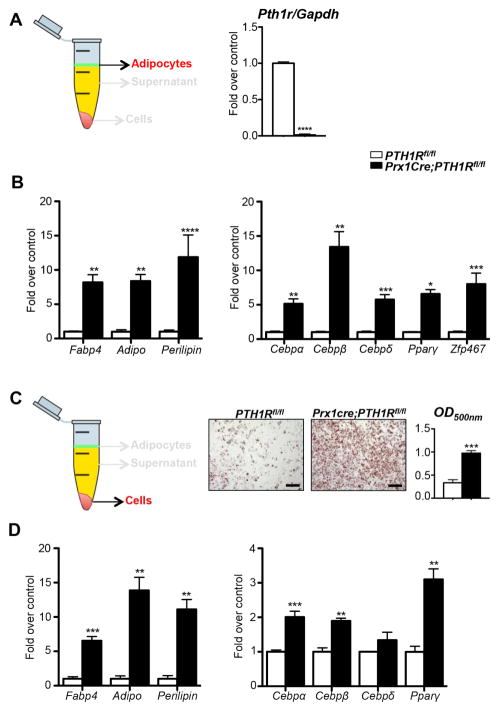
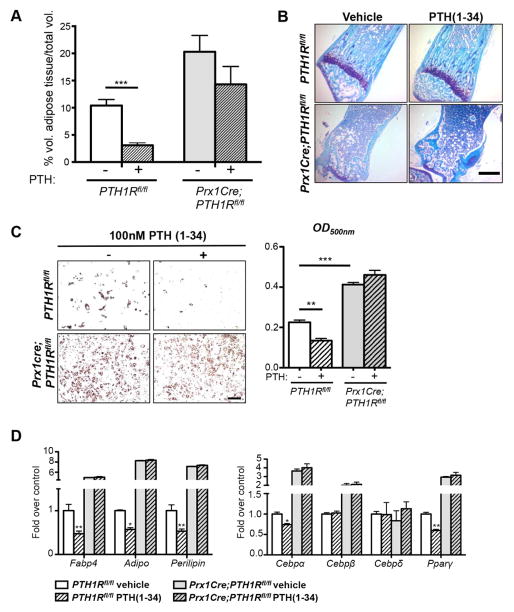
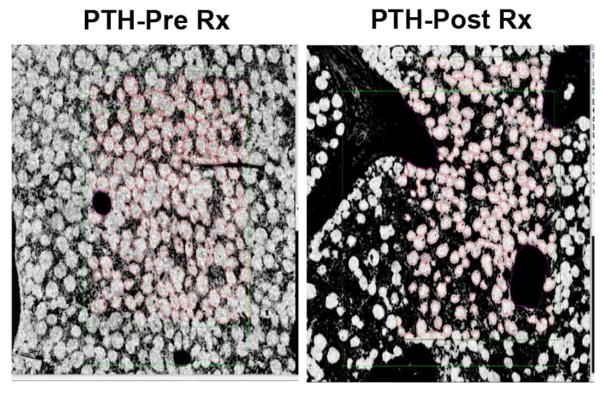
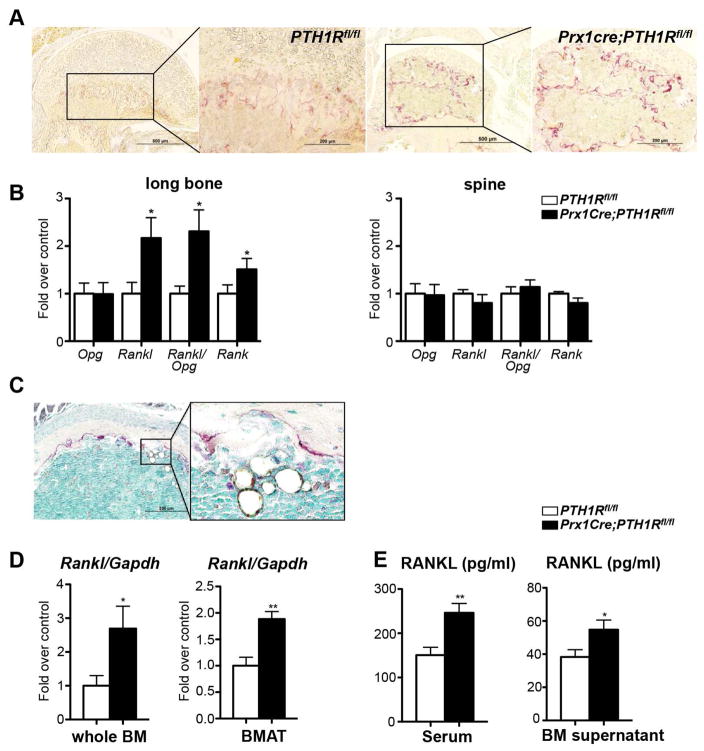
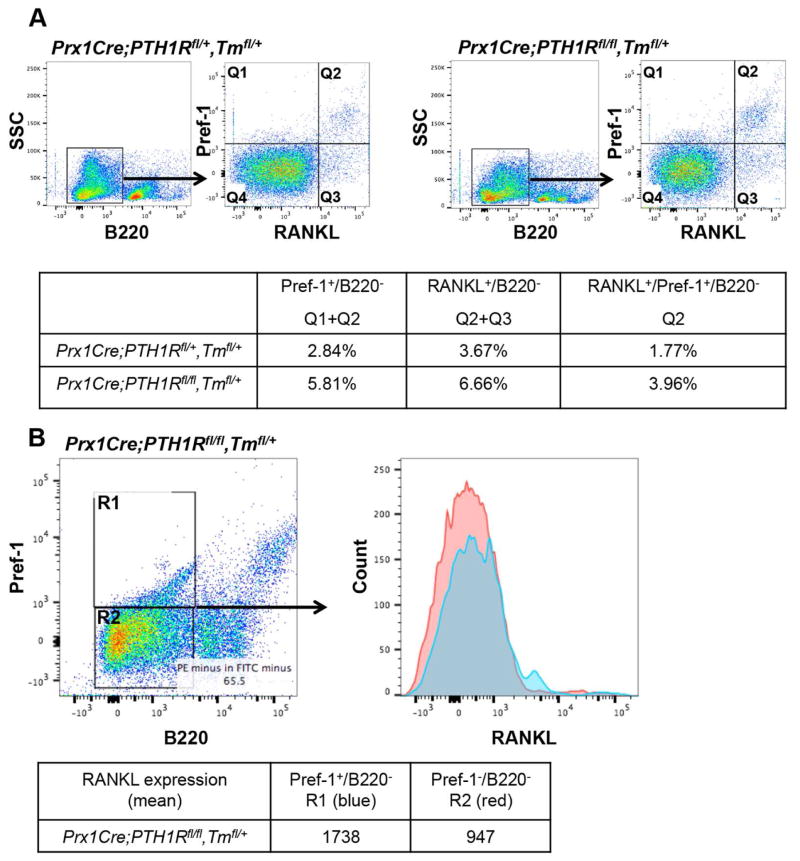
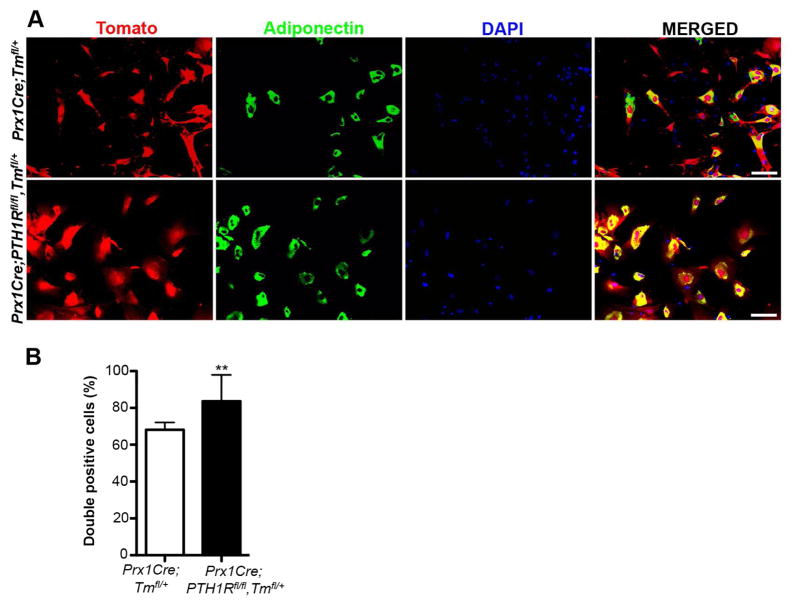
Intermittent PTH administration builds bone mass and prevents fractures, but its mechanism of action is unclear. We genetically deleted the PTH/PTHrP receptor (PTH1R) in mesenchymal stem cells using Prx1Cre and found low bone formation, increased bone resorption, and high bone marrow adipose tissue (BMAT). Bone marrow adipocytes traced to Prx1 and expressed classic adipogenic markers and high receptor activator of nuclear factor kappa B ligand (Rankl) expression. RANKL levels were also elevated in bone marrow supernatant and serum, but undetectable in other adipose depots. By cell sorting, Pref1+RANKL+ marrow progenitors were twice as great in mutant versus control marrow. Intermittent PTH administration to control mice reduced BMAT significantly. A similar finding was noted in male osteoporotic patients. Thus, marrow adipocytes exhibit osteogenic and adipogenic characteristics, are uniquely responsive to PTH, and secrete RANKL. These studies reveal an important mechanism for PTH's therapeutic action through its ability to direct mesenchymal cell fate.
Keywords: PTH; RANKL; bone resorption; lineage; receptor.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Stem cells: PTH regulates bone marrow progenitor fate.Nat Rev Endocrinol. 2017 Apr;13(4):190. doi: 10.1038/nrendo.2017.19. Epub 2017 Feb 17. Nat Rev Endocrinol. 2017. PMID: 28211514 No abstract available.
References
-
- Amizuka N, Karaplis AC, Henderson JE, Warshawsky H, Lipman ML, Matsuki Y, Ejiri S, Tanaka M, Izumi N, Ozawa H, et al. Haploinsufficiency of parathyroid hormone-related peptide (PTHrP) results in abnormal postnatal bone development. Dev Biol. 1996;175:166–176. - PubMed
-
- Calvi LM, Sims NA, Hunzelman JL, Knight MC, Giovannetti A, Saxton JM, Kronenberg HM, Baron R, Schipani E. Activated parathyroid hormone/parathyroid hormone-related protein receptor in osteoblastic cells differentially affects cortical and trabecular bone. J Clin Invest. 2001;107:277–286. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
