

Discovering Targets of Non-enzymatic Acylation by Thioester Reactivity Profiling
- PMID: 28163016
- PMCID: PMC5864104
- DOI: 10.1016/j.chembiol.2017.01.002
Discovering Targets of Non-enzymatic Acylation by Thioester Reactivity Profiling
Abstract
Non-enzymatic protein modification driven by thioester reactivity is thought to play a major role in the establishment of cellular lysine acylation. However, the specific protein targets of this process are largely unknown. Here we report an experimental strategy to investigate non-enzymatic acylation in cells. Specifically, we develop a chemoproteomic method that separates thioester reactivity from enzymatic utilization, allowing selective enrichment of non-enzymatic acylation targets. Applying this method to cancer cell lines identifies numerous candidate targets of non-enzymatic acylation, including several enzymes in lower glycolysis. Functional studies highlight malonyl-CoA as a reactive thioester metabolite that can modify and inhibit glycolytic enzyme activity. Finally, we show that synthetic thioesters can be used as novel reagents to probe non-enzymatic acylation in living cells. Our studies provide new insights into the targets and drivers of non-enzymatic acylation, and demonstrate the utility of reactivity-based methods to experimentally investigate this phenomenon in biology and disease.
Keywords: Warburg effect; acetylation; acylation; epigenetics; glycolysis; malonylation; metabolism; non-enzymatic; reactivity-based protein profiling; thioester.
Published by Elsevier Ltd.
Figures
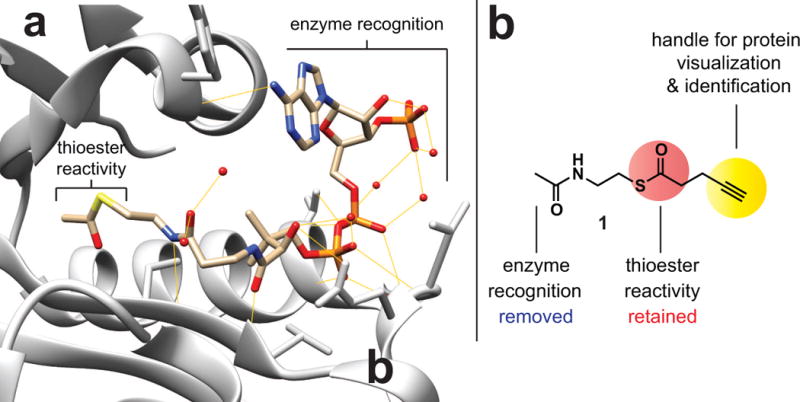

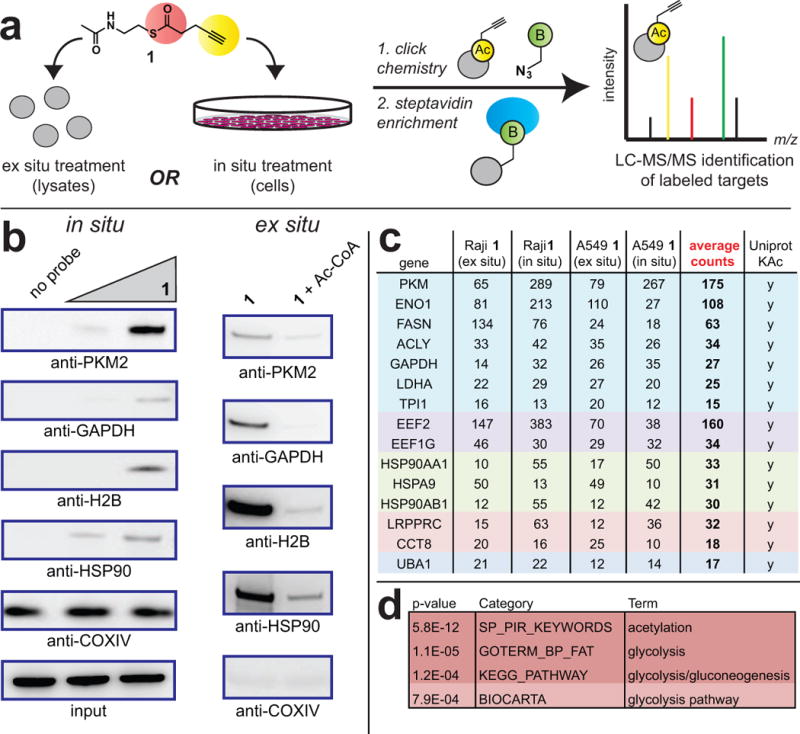
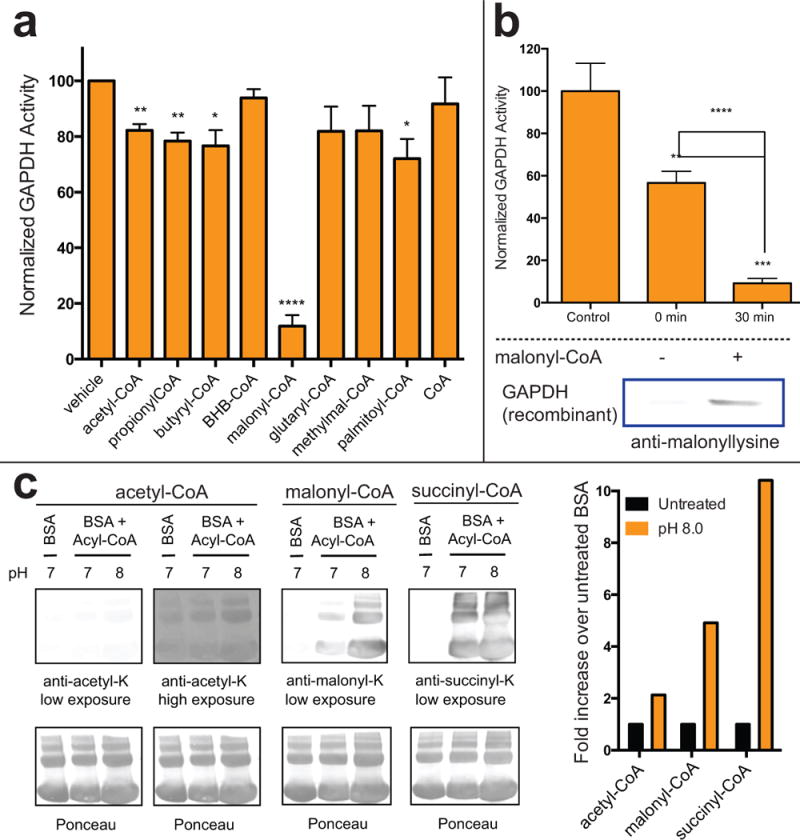

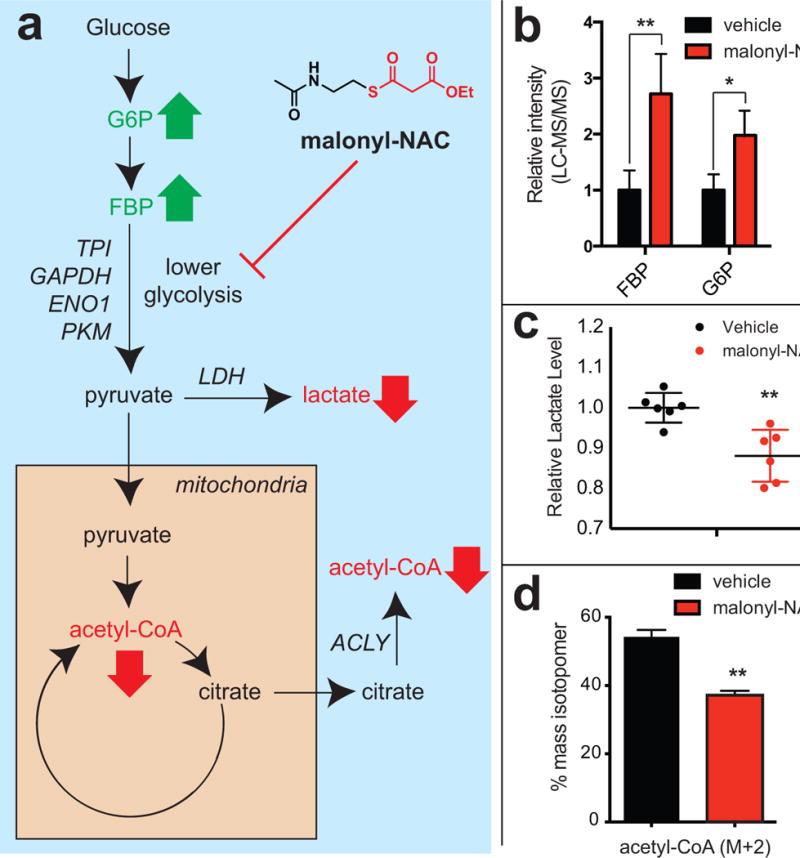
Comment in
-
A Prob(e)able Route to Lysine Acylation.Cell Chem Biol. 2017 Feb 16;24(2):126-128. doi: 10.1016/j.chembiol.2017.01.011. Cell Chem Biol. 2017. PMID: 28212757 Free PMC article.
References
-
- Candiano G, Bruschi M, Musante L, Santucci L, Ghiggeri GM, Carnemolla B, Orecchia P, Zardi L, Righetti PG. Blue silver: a very sensitive colloidal Coomassie G-250 staining for proteome analysis. Electrophoresis. 2004;25:1327–1333. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
