

Killing by Type VI secretion drives genetic phase separation and correlates with increased cooperation
- PMID: 28165005
- PMCID: PMC5303878
- DOI: 10.1038/ncomms14371
Killing by Type VI secretion drives genetic phase separation and correlates with increased cooperation
Abstract
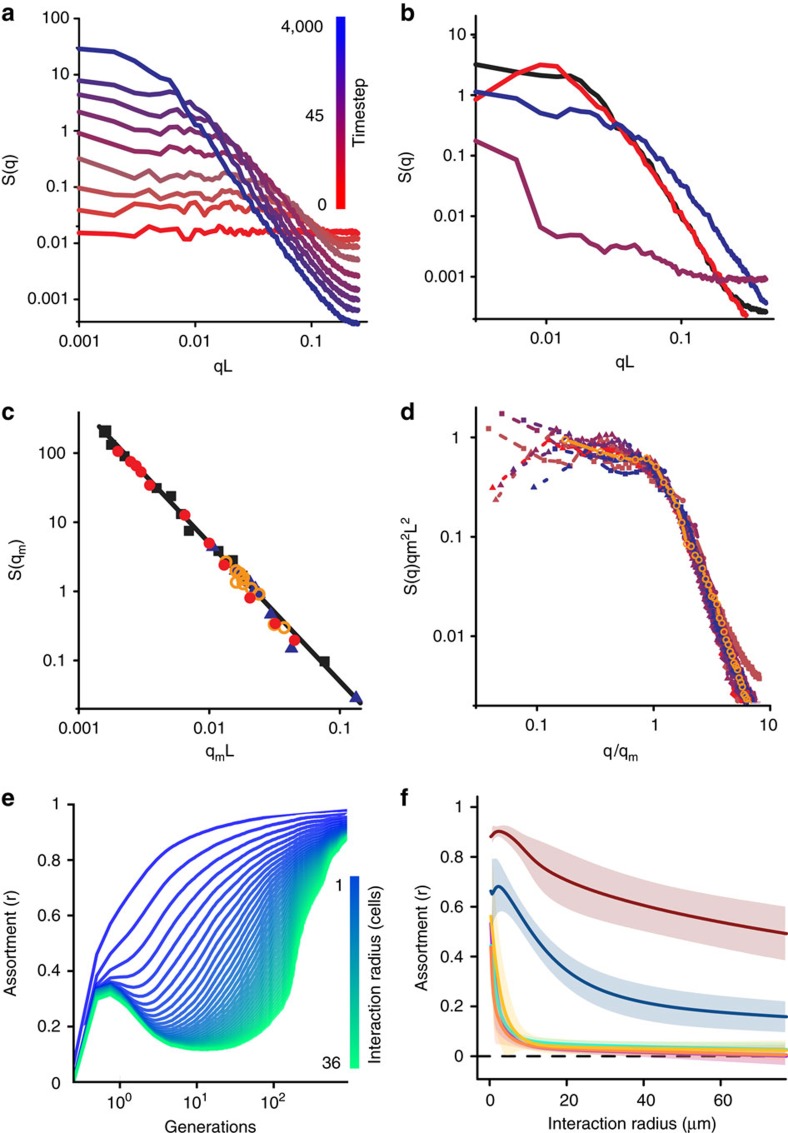
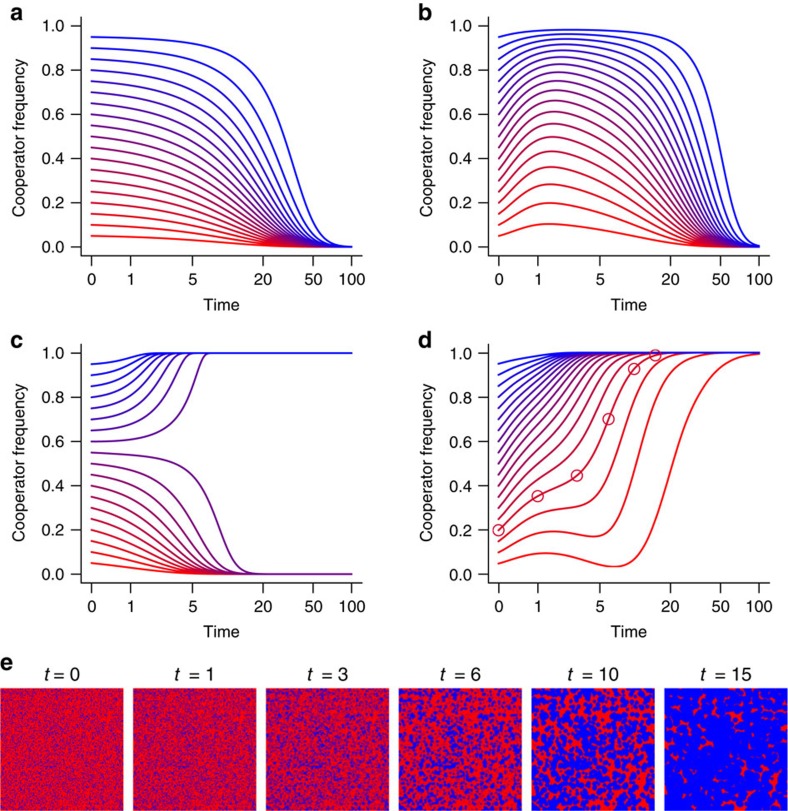
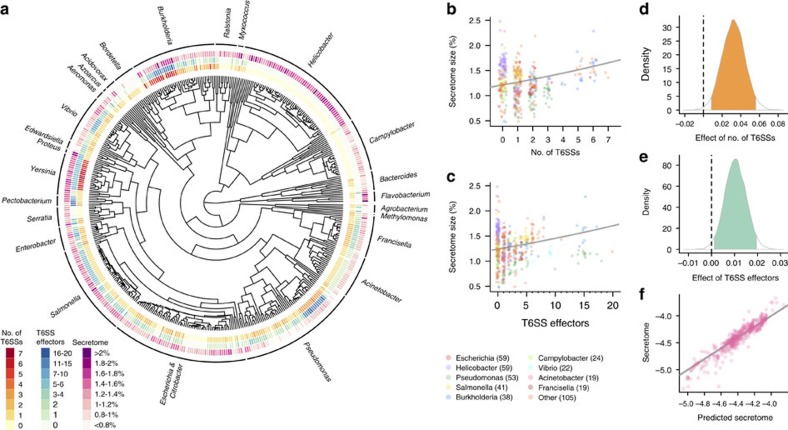
By nature of their small size, dense growth and frequent need for extracellular metabolism, microbes face persistent public goods dilemmas. Genetic assortment is the only general solution stabilizing cooperation, but all known mechanisms structuring microbial populations depend on the availability of free space, an often unrealistic constraint. Here we describe a class of self-organization that operates within densely packed bacterial populations. Through mathematical modelling and experiments with Vibrio cholerae, we show how killing adjacent competitors via the Type VI secretion system (T6SS) precipitates phase separation via the 'Model A' universality class of order-disorder transition mediated by killing. We mathematically demonstrate that T6SS-mediated killing should favour the evolution of public goods cooperation, and empirically support this prediction using a phylogenetic comparative analysis. This work illustrates the twin role played by the T6SS, dealing death to local competitors while simultaneously creating conditions potentially favouring the evolution of cooperation with kin.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

Similar articles
-
Horizontal Gene Transfer of Functional Type VI Killing Genes by Natural Transformation.mBio. 2017 Jul 25;8(4):e00654-17. doi: 10.1128/mBio.00654-17. mBio. 2017. PMID: 28743812 Free PMC article.
-
Quorum-sensing- and type VI secretion-mediated spatiotemporal cell death drives genetic diversity in Vibrio cholerae.Cell. 2022 Oct 13;185(21):3966-3979.e13. doi: 10.1016/j.cell.2022.09.003. Epub 2022 Sep 26. Cell. 2022. PMID: 36167071 Free PMC article.
-
Rules of Engagement: The Type VI Secretion System in Vibrio cholerae.Trends Microbiol. 2017 Apr;25(4):267-279. doi: 10.1016/j.tim.2016.12.003. Epub 2016 Dec 24. Trends Microbiol. 2017. PMID: 28027803 Free PMC article. Review.
-
Ecotin and LamB in Escherichia coli influence the susceptibility to Type VI secretion-mediated interbacterial competition and killing by Vibrio cholerae.Biochim Biophys Acta Gen Subj. 2021 Jul;1865(7):129912. doi: 10.1016/j.bbagen.2021.129912. Epub 2021 Apr 20. Biochim Biophys Acta Gen Subj. 2021. PMID: 33892013
-
Causalities of war: The connection between type VI secretion system and microbiota.Cell Microbiol. 2020 Mar;22(3):e13153. doi: 10.1111/cmi.13153. Epub 2020 Jan 12. Cell Microbiol. 2020. PMID: 31872954 Free PMC article. Review.
Cited by
-
Trojan horselike T6SS effector TepC mediates both interference competition and exploitative competition.ISME J. 2024 Jan 8;18(1):wrad028. doi: 10.1093/ismejo/wrad028. ISME J. 2024. PMID: 38365238 Free PMC article.
-
Biofilm Bacteria Use Stress Responses to Detect and Respond to Competitors.Curr Biol. 2020 Apr 6;30(7):1231-1244.e4. doi: 10.1016/j.cub.2020.01.065. Epub 2020 Feb 20. Curr Biol. 2020. PMID: 32084407 Free PMC article.
-
Biofilm formation and toxin production provide a fitness advantage in mixed colonies of environmental yeast isolates.Ecol Evol. 2018 Apr 27;8(11):5541-5550. doi: 10.1002/ece3.4082. eCollection 2018 Jun. Ecol Evol. 2018. PMID: 29938072 Free PMC article.
-
Mechanism of Kin-Discriminatory Demarcation Line Formation between Colonies of Swarming Bacteria.Biophys J. 2017 Dec 5;113(11):2477-2486. doi: 10.1016/j.bpj.2017.09.020. Biophys J. 2017. PMID: 29212001 Free PMC article.
-
Signatures of kin selection in a natural population of the bacteria Bacillus subtilis.Evol Lett. 2023 Jul 18;7(5):315-330. doi: 10.1093/evlett/qrad029. eCollection 2023 Oct. Evol Lett. 2023. PMID: 37829498 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
